Aβ Acts Through Pericytes to Throttle Brain Blood Flow
Quick Links
In Alzheimer’s disease, blood flows sluggishly through the brain, partly because Aβ provokes small blood vessels to constrict. In the June 20 Science, researchers led by David Attwell at University College London blame this on pericytes, tiny cells that wrap around capillaries. The researchers found that in rodent brain, synthetic Aβ42 oligomers sparked formation of reactive oxygen species in pericytes, leading to release of the vasoconstrictor endothelin-1. Endothelin-1 in turn instructed pericytes to squeeze the blood vessels, narrowing diameter by about a quarter.
- Aβ stimulates production of reactive oxygen species in cortical pericytes.
- In response, these cells tighten around capillaries, slowing blood flow.
- Targeting these cells might help restore vascular function.
In human AD brain tissue, too, pericytes appeared to pinch capillaries. This would have increased the overall vascular resistance threefold, and stemmed cerebral blood flow by about half, Attwell calculated. This is similar to the actual decrease in blood flow measured in AD (Asllani et al., 2008). “Provocatively, one can ask what fraction of the neuronal dysfunction and damage occurring in AD is caused directly by Aβ and tau, and what fraction reflects the decrease in energy supplied by the blood,” Attwell wrote to Alzforum. He believes therapeutics that restore vascular health could prove beneficial in AD.
Costantino Iadecola at Weill Cornell Medical College in New York agreed the findings extend researchers’ understanding of vascular dysfunction in Alzheimer’s. “[The data] point to an emerging role of cerebral capillaries in the vascular pathobiology of Aβ, and reinforce the idea that Aβ acts at all levels of the cerebrovascular tree and on different cells to exert deleterious effects on the brain circulation,” he wrote to Alzforum (full comment below). He noted that the next step will be to show that preventing pericyte vasoconstriction can rescue learning and memory in mice.
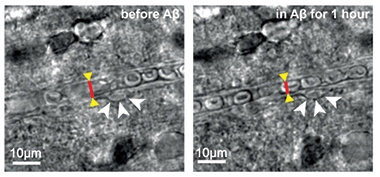
Restricted Flow. The width (red bar) of a rat cortical capillary (left) shrinks in the vicinity of a pericyte (white arrowheads) after addition of Aβ (right). [Courtesy of Nortley et al., Science/AAAS.]
Researchers have known for decades that Aβ40 and Aβ42 tighten brain blood vessels, and that this occurs through generation of reactive oxygen species (Suo et al., 1998; Niwa et al., 2001; Dietrich et al., 2010). Previous work from Iadecola blamed macrophages near arterioles for ROS production (Park et al., 2011; Park et al., 2017). Attwell wondered if pericytes and capillaries might be involved. While the role of pericytes in regulating blood flow is controversial (Feb 2017 news; May 2017 news), Attwell previously found that a subset of these cells contain α-smooth muscle actin and can squeeze capillaries, reducing their diameter (Peppiatt et al., 2006; Hall et al., 2014).
To see if this happens in response to Aβ, first author Ross Nortley applied synthetic oligomeric Aβ40 or Aβ42 to cortical brain slices taken from rats. An hour of exposure to either peptide narrowed capillaries near pericytes by 15 percent. The degree of narrowing depended on Aβ concentration, reaching its maximal effect at 20 nM Aβ42, and half-maximal at 5 nM. In AD brain, soluble Aβ has been measured at 6 nM, Attwell noted (Roberts et al., 2017).
Through what cell types in these slices did Aβ act? Aβ is known to prod NADPH oxidases to form reactive superoxides. The NADPH oxidase found in pericytes and endothelial cells is NOX4, and in immune cells, NOX2. A NOX4 blocker prevented capillary constriction after Aβ application, while a NOX2 blocker only partially ameliorated it. This suggested that pericytes play a larger role than macrophages in mediating Aβ’s effects, the authors concluded. Supporting this, Aβ exposure boosted ROS in cortical pericytes by sevenfold, but in macrophages only twofold.
ROS enhance transcription of the vasoconstrictor endothelin-1 in pericytes, microglia, and endothelial cells (Belaidi et al., 2016). Pericytes also express the endothelin-1 type A receptor. If the authors blocked this receptor as they added Aβ, pericytes did not clamp down on capillaries. Applying C-type natriuretic peptide (CNP) to rat cortical slices had a more powerful effect, relaxing capillaries that had previously tightened. This compound is known to reverse the effects of endothelin-1.
Do pericytes in human brain respond the same way? Nortley and colleagues obtained human cortical tissue that had been removed during neurosurgeries, and added synthetic oligomeric Aβ42 to these slices. Forty minutes of exposure shrank capillary diameter by 25 percent. To see if this narrowing was associated with pericytes, the authors examined postmortem cortical tissue from four people with a high amyloid plaque load, three with moderate plaque, and six age-matched controls without amyloid deposition. Though the number of samples was small, capillaries around pericytes were 30 percent narrower in amyloid-laden brain tissue than in control tissue. The degree of narrowing correlated with plaque load.
The authors made similar observations in APPNL-G-F knock-in mice. In four such mice, capillary diameter around cortical pericytes was down about a quarter compared to three wild-types, but there was no difference in capillaries in the cerebellum, where plaques are scarce.
Attwell noted that boosting cerebral blood flow may help ameliorate AD, particularly because low blood flow itself stimulates Aβ production (Sun et al., 2006; Zhang et al., 2007). He suggested several potential approaches, such as inhibiting NOX4, mopping up ROS, suppressing endothelin-1 release, or blocking its receptor. He is testing these approaches in APPNL-G-F mice.—Madolyn Bowman Rogers
References
News Citations
Research Models Citations
Paper Citations
- Asllani I, Habeck C, Scarmeas N, Borogovac A, Brown TR, Stern Y. Multivariate and univariate analysis of continuous arterial spin labeling perfusion MRI in Alzheimer's disease. J Cereb Blood Flow Metab. 2008 Apr;28(4):725-36. PubMed.
- Suo Z, Humphrey J, Kundtz A, Sethi F, Placzek A, Crawford F, Mullan M. Soluble Alzheimers beta-amyloid constricts the cerebral vasculature in vivo. Neurosci Lett. 1998 Nov 27;257(2):77-80. PubMed.
- Niwa K, Porter VA, Kazama K, Cornfield D, Carlson GA, Iadecola C. A beta-peptides enhance vasoconstriction in cerebral circulation. Am J Physiol Heart Circ Physiol. 2001 Dec;281(6):H2417-24. PubMed.
- Dietrich HH, Xiang C, Han BH, Zipfel GJ, Holtzman DM. Soluble amyloid-beta, effect on cerebral arteriolar regulation and vascular cells. Mol Neurodegener. 2010;5:15. PubMed.
- Park L, Wang G, Zhou P, Zhou J, Pitstick R, Previti ML, Younkin L, Younkin SG, Van Nostrand WE, Cho S, Anrather J, Carlson GA, Iadecola C. Scavenger receptor CD36 is essential for the cerebrovascular oxidative stress and neurovascular dysfunction induced by amyloid-beta. Proc Natl Acad Sci U S A. 2011 Mar 22;108(12):5063-8. PubMed.
- Park L, Uekawa K, Garcia-Bonilla L, Koizumi K, Murphy M, Pistik R, Younkin L, Younkin S, Zhou P, Carlson G, Anrather J, Iadecola C. Brain Perivascular Macrophages Initiate the Neurovascular Dysfunction of Alzheimer Aβ Peptides. Circ Res. 2017 Jul 21;121(3):258-269. Epub 2017 May 17 PubMed.
- Peppiatt CM, Howarth C, Mobbs P, Attwell D. Bidirectional control of CNS capillary diameter by pericytes. Nature. 2006 Oct 12;443(7112):700-4. Epub 2006 Oct 1 PubMed.
- Hall CN, Reynell C, Gesslein B, Hamilton NB, Mishra A, Sutherland BA, O'Farrell FM, Buchan AM, Lauritzen M, Attwell D. Capillary pericytes regulate cerebral blood flow in health and disease. Nature. 2014 Apr 3;508(7494):55-60. Epub 2014 Mar 26 PubMed.
- Roberts BR, Lind M, Wagen AZ, Rembach A, Frugier T, Li QX, Ryan TM, McLean CA, Doecke JD, Rowe CC, Villemagne VL, Masters CL. Biochemically-defined pools of amyloid-β in sporadic Alzheimer's disease: correlation with amyloid PET. Brain. 2017 May 1;140(5):1486-1498. PubMed.
- Belaidi E, Morand J, Gras E, Pépin JL, Godin-Ribuot D. Targeting the ROS-HIF-1-endothelin axis as a therapeutic approach for the treatment of obstructive sleep apnea-related cardiovascular complications. Pharmacol Ther. 2016 Dec;168:1-11. Epub 2016 Aug 2 PubMed.
- Sun X, He G, Qing H, Zhou W, Dobie F, Cai F, Staufenbiel M, Huang LE, Song W. Hypoxia facilitates Alzheimer's disease pathogenesis by up-regulating BACE1 gene expression. Proc Natl Acad Sci U S A. 2006 Dec 5;103(49):18727-32. PubMed.
- Zhang X, Zhou K, Wang R, Cui J, Lipton SA, Liao FF, Xu H, Zhang YW. Hypoxia-inducible factor 1alpha (HIF-1alpha)-mediated hypoxia increases BACE1 expression and beta-amyloid generation. J Biol Chem. 2007 Apr 13;282(15):10873-80. PubMed.
Further Reading
News
- Absent Aβ, Blood-Brain Barrier Breakdown Predicts Cognitive Impairment
- Loss of Pericytes Wreaks Havoc in Mouse Brain
- What’s Up With the Vasculature in Dementia?
- Smooth Muscle Cells, Not Pericytes, Control Brain Blood Flow
- Pericytes Don’t Go With the Flow—They Change It
- Finally, a Dye to Visualize Pericyte Function
Primary Papers
- Nortley R, Korte N, Izquierdo P, Hirunpattarasilp C, Mishra A, Jaunmuktane Z, Kyrargyri V, Pfeiffer T, Khennouf L, Madry C, Gong H, Richard-Loendt A, Huang W, Saito T, Saido TC, Brandner S, Sethi H, Attwell D. Amyloid β oligomers constrict human capillaries in Alzheimer's disease via signaling to pericytes. Science. 2019 Jul 19;365(6450) Epub 2019 Jun 20 PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
Weill College Medicine, New York
The story of the vascular effects of Aβ is evolving steadily, and the present paper, in conjunction with the paper of Cruz Hernández et al., 2019, represents the latest chapters.
It has been known for a while that Aβ is a vasoconstrictor acting through free radicals derived from NADPH oxidase, and that counteracting these vascular effects rescues cognitive impairment in mouse models of Aβ accumulation, attesting to their pathogenic relevance (Park et al., 2005; Park et al., 2008). Subsequent studies identified the Aβ-binding scavenger receptor CD36 and NOX2 in perivascular macrophage associated with resistance arterioles as the source of the radicals and contributing to the neurovascular dysfunction (Park et al., 2011; Park et al., 2017). The ultimate effector of the dysfunction, downstream of radicals, was identified to be dysregulated opening of the TRPM2 channels and Ca2+ overload in cerebral endothelial cells (Park et al., 2014).
The present study extends these observations in several respects. First, it implicates vasoactivity of capillaries in the microvascular effects of Aβ. Second, it identifies capillary mural cells, such as pericytes, as critical for the capillary constriction. Third, it suggests that in these cells NOX4, rather than NOX2, is the source of the radicals involved in the capillary constriction. Fourth, and most importantly, it verifies some of these observations in freshly collected human brain samples. The importance of capillaries as a relevant microvascular site for the effects of Aβ on cerebral perfusion is also suggested by the Cruz Hernández study, demonstrating that transient occlusions of capillaries by circulating leukocytes (capillary stalling) contributes to the cerebral hypoperfusion associated with Aβ accumulation, and that counteracting the stalling rescues the reduction of cerebral blood flow and cognitive function in mice overexpressing APP (Cruz Hernández et al., 2019).
These observations, in concert with accumulating evidence of blood-brain barrier dysfunction in AD (Nation et al., 2019) and capillary transit time abnormality (Gutiérrez-Jiménez et al., 2018), point to an emerging role of cerebral capillaries in the vascular pathobiology of Aβ and reinforce the idea that Aβ acts at all levels of the cerebrovascular tree and on different cells to exert its deleterious effects on the brain circulation. Evidence demonstrating that rescuing the pericyte contribution to this process ameliorates cognitive function would be the next step in building the case that targeting pericytes and capillary vasoactivity is a viable strategy to counteract the neurovascular dysfunction induced by Aβ.
References:
Cruz Hernández JC, Bracko O, Kersbergen CJ, Muse V, Haft-Javaherian M, Berg M, Park L, Vinarcsik LK, Ivasyk I, Rivera DA, Kang Y, Cortes-Canteli M, Peyrounette M, Doyeux V, Smith A, Zhou J, Otte G, Beverly JD, Davenport E, Davit Y, Lin CP, Strickland S, Iadecola C, Lorthois S, Nishimura N, Schaffer CB. Neutrophil adhesion in brain capillaries reduces cortical blood flow and impairs memory function in Alzheimer's disease mouse models. Nat Neurosci. 2019 Mar;22(3):413-420. Epub 2019 Feb 11 PubMed.
Park L, Anrather J, Zhou P, Frys K, Pitstick R, Younkin S, Carlson GA, Iadecola C. NADPH-oxidase-derived reactive oxygen species mediate the cerebrovascular dysfunction induced by the amyloid beta peptide. J Neurosci. 2005 Feb 16;25(7):1769-77. PubMed.
Park L, Zhou P, Pitstick R, Capone C, Anrather J, Norris EH, Younkin L, Younkin S, Carlson G, McEwen BS, Iadecola C. Nox2-derived radicals contribute to neurovascular and behavioral dysfunction in mice overexpressing the amyloid precursor protein. Proc Natl Acad Sci U S A. 2008 Jan 29;105(4):1347-52. PubMed.
Park L, Wang G, Zhou P, Zhou J, Pitstick R, Previti ML, Younkin L, Younkin SG, Van Nostrand WE, Cho S, Anrather J, Carlson GA, Iadecola C. Scavenger receptor CD36 is essential for the cerebrovascular oxidative stress and neurovascular dysfunction induced by amyloid-beta. Proc Natl Acad Sci U S A. 2011 Mar 22;108(12):5063-8. PubMed.
Park L, Uekawa K, Garcia-Bonilla L, Koizumi K, Murphy M, Pistik R, Younkin L, Younkin S, Zhou P, Carlson G, Anrather J, Iadecola C. Brain Perivascular Macrophages Initiate the Neurovascular Dysfunction of Alzheimer Aβ Peptides. Circ Res. 2017 Jul 21;121(3):258-269. Epub 2017 May 17 PubMed.
Park L, Wang G, Moore J, Girouard H, Zhou P, Anrather J, Iadecola C. The key role of transient receptor potential melastatin-2 channels in amyloid-β-induced neurovascular dysfunction. Nat Commun. 2014 Oct 29;5:5318. PubMed.
Nation DA, Sweeney MD, Montagne A, Sagare AP, D'Orazio LM, Pachicano M, Sepehrband F, Nelson AR, Buennagel DP, Harrington MG, Benzinger TL, Fagan AM, Ringman JM, Schneider LS, Morris JC, Chui HC, Law M, Toga AW, Zlokovic BV. Blood-brain barrier breakdown is an early biomarker of human cognitive dysfunction. Nat Med. 2019 Feb;25(2):270-276. Epub 2019 Jan 14 PubMed.
Gutiérrez-Jiménez E, Angleys H, Rasmussen PM, West MJ, Catalini L, Iversen NK, Jensen MS, Frische S, Østergaard L. Disturbances in the control of capillary flow in an aged APPswe /PS1ΔE9 model of Alzheimer's disease. Neurobiol Aging. 2018 Feb;62:82-94. Epub 2017 Oct 16 PubMed.
University of Southern California
University of Southern California, Keck School of Medicine
Vasoactive properties of peptides have been known for more than 20 years. For example, in the mid-1990s Aβ peptides were shown to induce vasoconstriction in the isolated rat aorta, which was ameliorated by superoxide dismutase 1 (SOD1) implicating involvement of reactive oxygen species (ROS) (Thomas et al., 1996). The follow-up studies in young Tg2576 mice overexpressing human APP (Swedish mutation) indicated that Aβ reduces cerebrovascular reactivity to endothelium-dependent vasodilators (for example, acetylcholine, bradykinin, or calcium ionophore A23187) and increases response to vasoconstrictors acting directly on vascular smooth muscle cells (VSMCs) (for example, the thromboxane A2 analogue U46619) (Zhang et al., 1997; Iadecola et al., 1999), that was associated with impaired neurovascular coupling (the dynamic functional change in cerebral blood flow that occurs in response to local neuronal activity) (Niwa et al., 2000). These abnormal vascular responses could again be rescued with SOD1, confirming involvement of ROS in vivo (Iadecola et al. 1999). Collectively, these pioneering studies suggest that accumulation of low levels of soluble Aβ (likely Aβ oligomeric species) before Aβ deposition leads to a global impairment of vascular responses. Moreover, in the brain endothelium Aβ-mediated oxidant stress led to proinflammatory changes in cerebral blood vessels and generation of endothelin 1 (ET1) through the receptor for advanced glycosylation end products (RAGE), which binds Aβ, that was shown to lead to cerebral blood flow (CBF) reductions in Tg2576 mice in vivo (Deane et al., 2003, 2012). Based on these findings (Deane et al., 2003), a clinical Phase 2/3 trial with a RAGE blocker in patients with mild Alzheimer’s disease and impaired glucose tolerance (NCT03980730 June 2019–July 2023) has been recently initiated.
Here, Nortley et al. importantly confirmed that Aβ generates ROS in rat brain slices, which evoked release of ET1 and activated ETa receptor in pericytes to cause capillary constriction, consistent with the proposed mechanism of ET1 activation of pericytes, as we recently reviewed (Kisler et al., 2017, see Figure 3). They extended studies to live healthy human brain slices, and human tissue derived from individuals with cognitive decline to show capillary constriction in areas rich with Aβ deposits close to PDGRFβ -positive pericytes, but not aSMA-positive arterioles. Interestingly, the authors only show that constriction occurs in capillaries in vivo, in contrast to their ex vivo data and previous reports implicating ROS-mediated vascular dysfunction in arterioles as well.
Although Nortley et al. propose that the ET1 pathway in pericytes could be a key mechanism implicated in neurodegeneration, they did not show any neuron and/or synaptic loss in their study, thus leaving open the question whether the proposed ET1-ETa mechanism can drive neurodegeneration. It has been reported, however, that accelerated pericyte loss in Tg2576 APP mice leads to both tau pathology and an overt neuron loss, suggesting that vascular damage caused by pericyte loss is a second hit in the disease process leading to neuron loss and degenerative changes in this AD model that otherwise does not occur (Sagare et al., 2013).
Other neuropathological studies in AD brain tissue have shown loss of pericytes associated with blood-brain barrier (BBB) breakdown and accumulation of Aβ deposits (Sengillo et al. 2013; Halliday et al. 2015). It has also been shown that Aβ intracellular accumulation kills pericytes (Ma et al., 2018; Sagare et al., 2013). Thus, while interesting, it remains to be seen how findings that Nortley et al. report fit with the existing knowledge in the field.
References:
Thomas T, Thomas G, McLendon C, Sutton T, Mullan M. beta-Amyloid-mediated vasoactivity and vascular endothelial damage. Nature. 1996 Mar 14;380(6570):168-71. PubMed.
Zhang F, Eckman C, Younkin S, Hsiao KK, Iadecola C. Increased susceptibility to ischemic brain damage in transgenic mice overexpressing the amyloid precursor protein. J Neurosci. 1997 Oct 15;17(20):7655-61. PubMed.
Iadecola C, Zhang F, Niwa K, Eckman C, Turner SK, Fischer E, Younkin S, Borchelt DR, Hsiao KK, Carlson GA. SOD1 rescues cerebral endothelial dysfunction in mice overexpressing amyloid precursor protein. Nat Neurosci. 1999 Feb;2(2):157-61. PubMed.
Niwa K, Younkin L, Ebeling C, Turner SK, Westaway D, Younkin S, Ashe KH, Carlson GA, Iadecola C. Abeta 1-40-related reduction in functional hyperemia in mouse neocortex during somatosensory activation. Proc Natl Acad Sci U S A. 2000 Aug 15;97(17):9735-40. PubMed.
Deane R, Du Yan S, Submamaryan RK, LaRue B, Jovanovic S, Hogg E, Welch D, Manness L, Lin C, Yu J, Zhu H, Ghiso J, Frangione B, Stern A, Schmidt AM, Armstrong DL, Arnold B, Liliensiek B, Nawroth P, Hofman F, Kindy M, Stern D, Zlokovic B. RAGE mediates amyloid-beta peptide transport across the blood-brain barrier and accumulation in brain. Nat Med. 2003 Jul;9(7):907-13. PubMed.
Deane R, Singh I, Sagare AP, Bell RD, Ross NT, LaRue B, Love R, Perry S, Paquette N, Deane RJ, Thiyagarajan M, Zarcone T, Fritz G, Friedman AE, Miller BL, Zlokovic BV. A multimodal RAGE-specific inhibitor reduces amyloid β-mediated brain disorder in a mouse model of Alzheimer disease. J Clin Invest. 2012 Apr 2;122(4):1377-92. PubMed.
Kisler K, Nelson AR, Montagne A, Zlokovic BV. Cerebral blood flow regulation and neurovascular dysfunction in Alzheimer disease. Nat Rev Neurosci. 2017 Jul;18(7):419-434. Epub 2017 May 18 PubMed.
Sagare AP, Bell RD, Zhao Z, Ma Q, Winkler EA, Ramanathan A, Zlokovic BV. Pericyte loss influences Alzheimer-like neurodegeneration in mice. Nat Commun. 2013;4:2932. PubMed. Correction. RETRACTED
Sengillo JD, Winkler EA, Walker CT, Sullivan JS, Johnson M, Zlokovic BV. Deficiency in Mural Vascular Cells Coincides with Blood-Brain Barrier Disruption in Alzheimer's Disease. Brain Pathol. 2012 Nov 5; PubMed.
Halliday MR, Rege SV, Ma Q, Zhao Z, Miller CA, Winkler EA, Zlokovic BV. Accelerated pericyte degeneration and blood-brain barrier breakdown in apolipoprotein E4 carriers with Alzheimer's disease. J Cereb Blood Flow Metab. 2015 Mar 11; PubMed.
Ma Q, Zhao Z, Sagare AP, Wu Y, Wang M, Owens NC, Verghese PB, Herz J, Holtzman DM, Zlokovic BV. Blood-brain barrier-associated pericytes internalize and clear aggregated amyloid-β42 by LRP1-dependent apolipoprotein E isoform-specific mechanism. Mol Neurodegener. 2018 Oct 19;13(1):57. PubMed. Correction. Correction.
Sagare AP, Bell RD, Zhao Z, Ma Q, Winkler EA, Ramanathan A, Zlokovic BV. Pericyte loss influences Alzheimer-like neurodegeneration in mice. Nat Commun. 2013;4:2932. PubMed. Correction. RETRACTED
Picower Institute of MIT
Icahn School of Medicine at Mt. Sinai
Vascular abnormalities have been observed in dementia since the seminal studies of Alois Alzheimer and Oscar Fisher in the early 1900s. However, vascular abnormalities have often taken a back seat to the more prominent pathologies of amyloid plaques and neurofibrillary tangles. Yet, from numerous studies it is emerging that vascular dysfunction likely has a causal role neurodegeneration and dementia (Nation et al., 2019). Recent findings from David Attwell’s laboratory underscore this point and implicate pericytes in cerebral vascular constrictions that likely contribute to cognitive impairment.
Nortley and colleagues provide compelling evidence that in humans elevated Aβ induces pericytes to constrict brain microvasculature leading to ischemic brain regions which could contribute to neuronal damage and synaptic loss in AD. This fits into emerging evidence that vascular dysfunction has a prominent role in neurodegeneration and in particular targeting pericytes may be a viable strategy to counteract cognitive impairments induced by neurovascular dysfunction (Bell et al., 2012; Halliday et al., 2015).
Conversely as the authors note, this study suggests that pericytes may also have a role in cognitive resilience. Numerous aged humans accumulate high levels of Aβ but seemingly do not develop cognitive impairments. It is interesting to speculate that pericytes could play a role whereby among human genetic diversity there are individuals endowed with “protective” pericytes that do not constrict in response to Aβ or are prone to compensatory vasodilation. Delving into pericyte biology and how human genetic diversity influences their contributions to pathophysiology certainly promises to be an exciting future direction.
References:
Bell RD, Winkler EA, Singh I, Sagare AP, Deane R, Wu Z, Holtzman DM, Betsholtz C, Armulik A, Sallstrom J, Berk BC, Zlokovic BV. Apolipoprotein E controls cerebrovascular integrity via cyclophilin A. Nature. 2012 May 24;485(7399):512-6. PubMed. Correction.
Halliday MR, Rege SV, Ma Q, Zhao Z, Miller CA, Winkler EA, Zlokovic BV. Accelerated pericyte degeneration and blood-brain barrier breakdown in apolipoprotein E4 carriers with Alzheimer's disease. J Cereb Blood Flow Metab. 2015 Mar 11; PubMed.
Nation DA, Sweeney MD, Montagne A, Sagare AP, D'Orazio LM, Pachicano M, Sepehrband F, Nelson AR, Buennagel DP, Harrington MG, Benzinger TL, Fagan AM, Ringman JM, Schneider LS, Morris JC, Chui HC, Law M, Toga AW, Zlokovic BV. Blood-brain barrier breakdown is an early biomarker of human cognitive dysfunction. Nat Med. 2019 Feb;25(2):270-276. Epub 2019 Jan 14 PubMed.
CAST, Center for Advanced Studies and Technologynal Medicine University “G. d’Annunzio” Chieti-Pescara
The missing link. This is a must-read article on the convergence between the amyloid dysmetabolism and vascular dysfunctions.
Make a Comment
To make a comment you must login or register.