ALS Protein Said to Liquefy, Then Freeze en Route to Disease
Quick Links
The neurodegeneration-linked protein FUS undergoes phase transitions just the way matter can occur as gas, liquid, or solid, according to a paper in the August 27 Cell. And just like oil is healthier than butter, FUS’s liquid droplet form is better for the cell than its more condensed alter ego, propose senior authors Tony Hyman and Simon Alberti of the Max Planck Institute of Molecular Cell Biology and Genetics in Dresden, Germany. Consolidating from soluble molecules into liquid droplets allows FUS to structure transient organelles such as stress granules. While this is a physiological process, it also creates a precarious state. When FUS condenses further, it can harden into the solid aggregates associated with neurodegeneration, the authors suggest. Hyman and Alberti predict that other proteins implicated in neurodegeneration, such as tau or α-synuclein, might undergo similar phase transitions. Their work suggests a reason why certain proteins contain aggregation-prone domains—they might need them to form the liquid state.
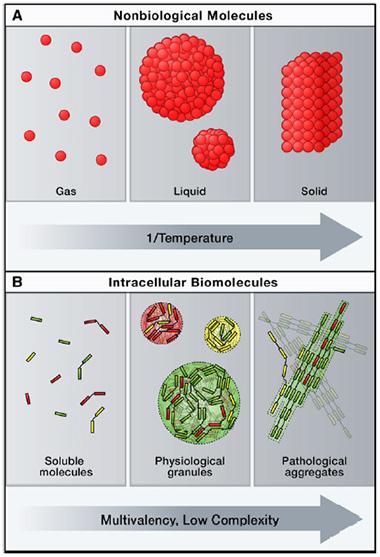
Phase transition.
Just as water condenses from vapor to liquid to ice, proteins can condense from a soluble phase to concentrated liquid “granules,” and further into solidified aggregates. [Courtesy of Cell, Weber and Brangwynne.]
Protein Liquefaction
It is known that proteins can undergo phase transitions akin to those of non-biological molecules (reviewed in Weber and Brangwynne, 2012; see image at right). Individual soluble proteins float through the cytosol like gas particles. Certain proteins, such as FUS, contain disordered, prion-like domains loaded with the sticky amino acids glutamine and asparagine. If these proteins become concentrated, they condense, like water vapor forming morning dew, explained Cliff Brangwynne of Princeton University in New Jersey, who did not participate in the current report. Cells use such proteins to create organelles unbound by membranes, such as the nucleolus and stress granules. While the organelles are distinct, their contents move constantly back and forth across the phase boundary, like the molecules of a water droplet in a humid room. Hyman compared them to the lipid rafts that form in membranes, except that these droplets are three-dimensional.
However, there is a third phase of matter. Proteins can solidify into crystals or gels. “The molecules all of a sudden snap into ordered configurations,” Brangwynne said. At this point, few of the "frozen" proteins transition back and forth across the phase boundary. “If you start to irreversibly freeze things … this is something that is going to be pathological,” he said.
Interest in the phase transitions of proteins involved in organelle structure and neurodegeneration has been building for some time, commented Paul Taylor of St. Jude Children’s Research Hospital in Memphis, Tennessee, who did not contribute to the Cell study. About a decade ago, scientists noticed that prion-like sequences promote formation of stress granules and processing bodies, i.e., cytoplasmic organelles that hold inactive messenger RNAs (Gilks et al., 2004; Decker et al., 2007). In 2009, Brangwynne and Hyman reported that RNA-associated granules in nematode embryos behave like liquids, “evaporating” and recondensing (Brangwynne et al., 2009). Physically, they are not granules, Hyman said. “They have all the properties of a liquid drop.”
Later scientists suggested phase transitions could be a general mechanism to form a variety of organelles (Li et al., 2012). In 2012, researchers proposed a similar idea, that proteins with prion-like domains make fibrils, which form hydrogels that structure granular organelles (Kato et al., 2012). For his part, Taylor’s lab discovered that two other genes involved in ALS and multisystem proteinopathy, hnRNPA1 and hnRNPA2B1, encode proteins that assemble into liquid droplets (see Nov 2014 conference news).
Freezing FUS
In the Cell paper, first authors Avinash Patel and Kate Lee explored how the RNA-binding protein FUS might undergo phase transitions. Mutations in FUS cause ALS, and the protein aggregates in most forms of that disease as well as a subset of frontotemporal dementia. Normally, FUS assembles alongside damaged DNA and around mRNAs in nuclear or cytoplasmic granules (see Sep 2013 news; Dec 2013 news). To check if FUS naturally liquefied, Lee expressed a fluorescently tagged version, under its normal promoter, in HeLa and embryonic stem cells. As others have reported, it assembled into foci in the nucleus and cytoplasm—but were they liquid foci?
The authors reasoned that liquid FUS should meet three criteria: (1) The FUS molecules ought to move freely within the droplet; (2) They should be round; (3) Two droplets should be able to fuse into a new, spherical droplet. Lee tested the first requirement by photobleaching half of a FUS focus, and watching as the remaining fluorescent protein rapidly redistributed across the whole droplet. For the second, she used a digital-scanned, light-sheet microscope with structured illumination to image the FUS blobs in three dimensions, and they indeed formed neat spheres. With the same technique, she tracked individual granules, and watched them fuse together into new spheres, passing the third test. “The physiological role of these proteins is to form liquid drops,” Hyman concluded.
What allows the droplets to quickly coalesce and disappear as needed by the cell? The answer lies in the high concentration of FUS, Alberti said. Study co-author Marco Hein, in the laboratory of Matthias Mann at the Max Planck Institute of Biochemistry in Martinsried, Germany, used mass spectrometry to quantify FUS in HeLa cells. He estimated its concentration at two micromolars in the cytoplasm, and up to eight micromolars in the nucleus, placing it in the top 5 percent of cellular proteins ranked by abundance. This is a supersaturated concentration, and keeps FUS molecules right on the edge of liquefaction, Alberti said. “They easily flip from a diffuse soluble state into this liquid state.”
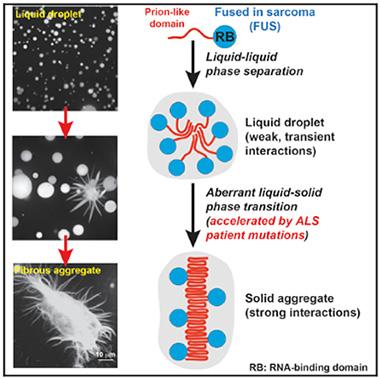
Liquid FUSion.
FUS, by virtue of its disordered prion-like domain, condenses into liquid droplets and solid aggregates. [Image courtesy of Cell, Patel et al.]
In order to liquefy, FUS also needs its prion-like domain. Patel discovered this when he used recombinant FUS protein to make the spherical blobs in test tubes. A version missing the sticky prion-like segment did not separate into droplets. Next, he tried to make droplets with FUS marred by an ALS-linked glycine-156-glutamate substitution in the prion-like domain. This mutant FUS made liquid droplets just fine in vitro, and in vivo as well.
People with FUS mutations do not develop ALS until late in life, suggesting it takes time for the pathology to arise. To mimic aging, the scientists let FUS drops sit in suspension for several hours. Over the first two hours, the mutant droplets fused only slowly, as if they were solidifying. By eight hours, they no longer fused at all. The wild-type FUS blobs also lost the ability to fuse, but not until 12 hours had passed.
As they stopped fusing, the droplets changed shape, from round to spiky and fibrous (see image above). This happened faster for both the G156E and another disease-linked mutation next to the prion-like domain, arginine-244-cysteine. “They convert from this lovely liquid drop to this horrible aggregate,” Hyman said. In other words, they froze.
Liquid Good, Solid Bad
The authors propose that FUS normally makes liquid droplets in the cell, allowing rapid assembly and disassembly of transient organelles. They believe the same disordered sequences that allow FUS to structure the liquid organelles make it prone to aggregate. Mutations promote aggregation by two possible mechanisms, they suggest. Changes in the prion-like domain may speed up nucleation of the solid state. Other ALS mutations in the nuclear localization sequence boost the cytosolic concentration of FUS and thus could also push it toward condensing.
“I think the paper is a real tour de force. It helps us understand why the prion-like domain of FUS is so useful (for forming functional liquids) but simultaneously so dangerous (as the liquids can turn into deleterious fibrils),” commented James Shorter of the Perelman School of Medicine at the University of Pennsylvania in Philadelphia, who was not involved in the work.
Brangwynne noted the authors do not report the conversion to solid FUS happening in cells, but he liked the theory. “The idea of physiological liquids and pathological solids makes a lot of sense,” he said.
Taylor agreed that formation of the liquid state, rather than the solid fibrils, likely underlies the birth of cellular granules. However, he thought fibrils might still contribute to granules without being pathological, for example by stabilizing the structure. Only when that process got out of control would pathological aggregates arise, he theorized.
Hyman and Alberti reason that the cell must have some way of keeping FUS liquid, perhaps a mechanism that falters with aging. They are pursuing this protective mechanism while also hunting for other proteins that undergo similar phase transitions. Both scientists believe, as does Brangwynne, that many proteins known to aggregate during disease may get there via a useful liquid state.—Amber Dance
References
News Citations
- Stream of Genetics Pushes FTD Research Forward
- Paper Alert: FUS a Fixer of Damaged DNA
- FUS RNA Granules Not So Stressed Out?
Paper Citations
- Weber SC, Brangwynne CP. Getting RNA and protein in phase. Cell. 2012 Jun 8;149(6):1188-91. PubMed.
- Gilks N, Kedersha N, Ayodele M, Shen L, Stoecklin G, Dember LM, Anderson P. Stress granule assembly is mediated by prion-like aggregation of TIA-1. Mol Biol Cell. 2004 Dec;15(12):5383-98. Epub 2004 Sep 15 PubMed.
- Decker CJ, Teixeira D, Parker R. Edc3p and a glutamine/asparagine-rich domain of Lsm4p function in processing body assembly in Saccharomyces cerevisiae. J Cell Biol. 2007 Nov 5;179(3):437-49. PubMed.
- Brangwynne CP, Eckmann CR, Courson DS, Rybarska A, Hoege C, Gharakhani J, Jülicher F, Hyman AA. Germline P granules are liquid droplets that localize by controlled dissolution/condensation. Science. 2009 Jun 26;324(5935):1729-32. Epub 2009 May 21 PubMed.
- Li P, Banjade S, Cheng HC, Kim S, Chen B, Guo L, Llaguno M, Hollingsworth JV, King DS, Banani SF, Russo PS, Jiang QX, Nixon BT, Rosen MK. Phase transitions in the assembly of multivalent signalling proteins. Nature. 2012 Mar 7;483(7389):336-40. PubMed.
- Kato M, Han TW, Xie S, Shi K, Du X, Wu LC, Mirzaei H, Goldsmith EJ, Longgood J, Pei J, Grishin NV, Frantz DE, Schneider JW, Chen S, Li L, Sawaya MR, Eisenberg D, Tycko R, McKnight SL. Cell-free formation of RNA granules: low complexity sequence domains form dynamic fibers within hydrogels. Cell. 2012 May 11;149(4):753-67. PubMed.
Further Reading
Papers
- Becker LA, Gitler AD. It's all starting to come together. Elife. 2015 Aug 5;4 PubMed.
- Toretsky JA, Wright PE. Assemblages: functional units formed by cellular phase separation. J Cell Biol. 2014 Sep 1;206(5):579-88. PubMed.
- Malinovska L, Kroschwald S, Alberti S. Protein disorder, prion propensities, and self-organizing macromolecular collectives. Biochim Biophys Acta. 2013 May;1834(5):918-31. Epub 2013 Jan 14 PubMed.
- Lee CF, Brangwynne CP, Gharakhani J, Hyman AA, Jülicher F. Spatial organization of the cell cytoplasm by position-dependent phase separation. Phys Rev Lett. 2013 Aug 23;111(8):088101. Epub 2013 Aug 20 PubMed.
- Brangwynne CP, Mitchison TJ, Hyman AA. Active liquid-like behavior of nucleoli determines their size and shape in Xenopus laevis oocytes. Proc Natl Acad Sci U S A. 2011 Mar 15;108(11):4334-9. Epub 2011 Feb 28 PubMed.
- Han TW, Kato M, Xie S, Wu LC, Mirzaei H, Pei J, Chen M, Xie Y, Allen J, Xiao G, McKnight SL. Cell-free formation of RNA granules: bound RNAs identify features and components of cellular assemblies. Cell. 2012 May 11;149(4):768-79. PubMed.
- Kedersha N, Ivanov P, Anderson P. Stress granules and cell signaling: more than just a passing phase?. Trends Biochem Sci. 2013 Oct;38(10):494-506. Epub 2013 Sep 10 PubMed.
Primary Papers
- Patel A, Lee HO, Jawerth L, Maharana S, Jahnel M, Hein MY, Stoynov S, Mahamid J, Saha S, Franzmann TM, Pozniakovski A, Poser I, Maghelli N, Royer LA, Weigert M, Myers EW, Grill S, Drechsel D, Hyman AA, Alberti S. A Liquid-to-Solid Phase Transition of the ALS Protein FUS Accelerated by Disease Mutation. Cell. 2015 Aug 27;162(5):1066-77. PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
Boston University School of Medicine
Increasing attention has focused on the liquid droplet nature of RNA-binding proteins. This physico-chemical characteristic has been well-known by RNA biologists for decades. However, the role of liquid gel transitions has come to the fore in neurodegeneration research because it could well play an important role in the pathophysiology of motor neuron diseases, and possibly other neurodegenerative diseases.
In this elegant paper, a team led by Hyman and Alberti takes this liquid gel transition model and shows that disease-linked mutations in FUS accelerate the transition to an aggregated solid state, which can then become persistent and pathological.
The team also highlights the role of PARPs in the phase transition. This adds an important new dimension to the approaches we need to use when evaluating these systems. This work provides a nice mixture of physical/chemical as well as cell biology to the field of neurodegenerative diseases, and is well done.
Make a Comment
To make a comment you must login or register.