Aβ Plaques: Breeding Ground for Toxic Tau?
Quick Links
In Alzheimer’s disease, Aβ plaques and aggregates of tau show up at different times in different regions of the brain. How the two pathologies interact remains unclear. Now, in the December 4 Nature Medicine, researchers led by Virginia Lee at the University of Pennsylvania, Philadelphia, report that neuritic plaques trigger the formation of a specific type of tau aggregate. These amyloid-associated tau clumps appear to then fuel the formation and spreading of other tau conglomerates, including neurofibrillary tangles and neuropil threads. The findings suggest that not only are Aβ plaques fertile grounds for initiating tau pathology, but that certain tau aggregates can seed other types, in different subcellular locations.
- In mice, neuritic plaques spawn a specific form of tau aggregate.
- NP tau aggregates faster and spreads more widely than other tau aggregates.
- NP tau may seed neurofibrillary tangles and neuropil threads.
“This is an exciting paper that nicely explores how Aβ interacts to enhance tau pathology,” wrote Rachel Bennett, who studies Aβ and tau interactions in Bradley Hyman’s lab at Massachusetts General Hospital in Charlestown. “[It shows] that Aβ can spatially alter tau pathology, shifting the balance toward a neuritic dystrophy or neurofibrillary tangle tau, depending on disease stage.” Takaomi Saido, RIKEN Brain Science Institute, Wako, Japan, found the work convincing. “The effort to understand the Aβ-tau axis has moved one step further,” he wrote (see full comment below).
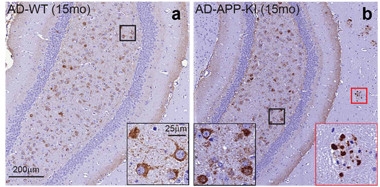
Plaque Forged Tau Aggregates.
Brains from wild-type (left) and APP-KI mice (right) develop neurofibrillary tangles (highlighted in black box) after injection of tau extracts from AD brains. Only APP-KI brains develop neuritic plaque tau aggregates that amplify and spread tau pathology (red box). [Courtesy of He et al., Nature Neuroscience.]
Researchers have long suspected that Aβ played a hand in turning tau toxic. In 1992, the amyloid cascade hypothesis proposed a link between Aβ and the formation of neurofibrillary tangles (NFTs) (Hardy and Higgins, 1992). Since then, researchers have reported enhanced formation of NFT-like aggregates after injecting Aβ fibrils into transgenic mice overexpressing mutant tau, and also in mice expressing both tau and Aβ precursor protein (APP) mutants (Aug 2001 news). More recently, Bennet, Hyman, and others reported that amyloid deposition in the cortices of APP/PS1-rTg4510 mice, which overexpress mutants of tau, APP, and presenilin1, accelerated and expanded tau propagation, with Aβ spurring tau to form specific pathological forms (Pooler et al., 2015; Bennett et al., 2017).
However, the mechanisms by which Aβ plaques fuel tau pathology remain uncertain. To gain a clearer view, first author Zhuohao He extracted a detergent-insoluble fraction enriched in tau paired-helical filaments, the major structural elements of NFTs, from two AD brains with abundant tau pathology (Guo et al., 2016). He then injected 2 μg of this AD-tau into the dorsal hippocampi and overlying cortices of 15-month-old wild-type mice and into two-, four-, and eight-month-old 5xFAD mice, as well as six- and 15-month-old APP knock-in mice (APP-KI). The 5xFAD mice have occasional and abundant Aβ plaques throughout the brain at two and eight months, respectively, while the APP-KI mice begin accumulating Aβ plaques in the cortex and hippocampus at six months, showing abundant plaques by 15 months. Researchers commended the use of these injection models because they avoided overexpression of mutant tau. “People have argued the frontotemporal dementia tau mutations used in other models produces non-AD-like tangles. This is a better, more natural model,” said Robert Vassar, Northwestern University, Evanston, Illinois.
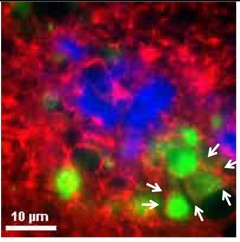
Neuritic tau.
Tau (green) accumulated within axons (red), adjacent to neuritic plaques (blue). [Courtesy of He et al., Nature Neuroscience.]
The authors found that wild-type and APP-KI mice injected with AD-tau at 15 months developed NFT-like aggregates three months later, as revealed by the anti-phospho-tau antibodies AT8 and AT180. The APP-KI mice also developed AT8-positive tau aggregates within dystrophic neurites adjacent to Aβ plaques (see image at left).
The 5xFAD mice told a different story. When the researchers injected AD-tau into eight-month-old animals they saw almost exclusively AT8-positive neuritic plaque tau (NP tau) three months later. “We were puzzled by this. Why didn’t we see NFTs?” said Lee. Repeating the experiment in younger, two-month-old 5xFAD mice, which have much lower Aβ plaque load, they saw both NFTs and NP tau. The data suggested that a high plaque load favored NP tau over NFTs, but the connection between the two tau pathologies remained unclear.
To gain more insight into the emergence of the different tau pathologies, the researchers then examined NFTs, NP tau, and a third type of aggregate, neuropil threads (NTs), at one, three, and six months post-injection. They used older mice: 15-month-old wild-type, 15-month-old APP-KI, and eight-month-old 5xFAD mice. NP tau accumulated first in APP-KI and 5xFAD mouse brains, spreading more quickly and more widely than NFTs. Wild-type mice developed no NP tau at all (see image below).
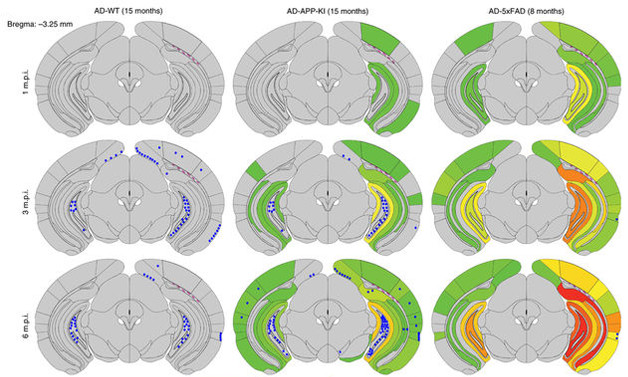
Aβ Plaques Fuel Tau Pathology. NP tau (green-red scale) accumulates faster and spreads more widely than NFTs (blue dots) in APP-KI (middle) and 5xFAD (right) mouse brains. Wild-type mice (left) develop no NP tau. [Courtesy of He et al., Nature Neuroscience.]
Putting all the data together, the researchers concluded that Aβ plaques likely trigger NP tau aggregates rather than NFTs. This would explain why wild-type mice develop only tangles and why Aβ-laden mice primarily form NP tau aggregates shortly after injection.
The researchers wondered if seeding of NFTs, which form in the soma, might be suppressed in AD models because the AD-tau gets caught up in the dystrophic neurites. In that case, adding more AD-tau might allow seeds to skirt the neurites and access the soma. Consistent with this idea, they found that by injecting 8 μg of AD-tau into four-month-old 5xFAD mice, which have less amyloid burden than the eight-month-old animals, they induced about four times the number of NFTs as when they used 2 μg. Still, even amyloid-laden mice injected with 2 μg of AD-tau eventually developed NFTs. In fact, while the number of NFTs plateaued after three months in wild-type mice, tangles continued to accumulate in 5xFAD for nine months, long after AD-tau had degraded. Lee and colleagues hypothesized that NP tau might migrate slowly from dystrophic neurites to seed NFTs in the soma and dendrites.
To test if NP tau can seed, the researchers extracted it from the hippocampi of 11-month-old 5xFAD mice that had been injected with AD-tau six months earlier, and injected it into the brains of wild-type mice. NFTs surfaced in multiple brain regions three months later. Neuropil threads also formed. The findings suggest that NP tau acts as a secondary tau seed that spreads tau pathology to different subcellular domains.
The researchers found evidence for a similar pattern in people. They stained postmortem brain sections with PHF-1, an antibody against tau paired-helical filaments, and with NAB228, an anti-Aβ antibody. They observed only NP tau in AD postmortem sections of the visual cortex at early stages of neurofibrillary pathology, but all three types of tau aggregates—NP tau, NFTs, and NTs—later on. The researchers focused on the visual cortex, one of the last brain regions to develop tau pathology in AD, to have a better chance of catching the early steps of the process.
What makes neuritic plaques such good breeding grounds for tau pathology? Endogenous mouse tau disengages from microtubules in the dystrophic axons surrounding Aβ plaques, the authors thought, becoming primed to interact with AD-tau seeds. In support of this, they found that in control eight-month-old 5xFAD mice, the pan-tau antibody, K9JA, bound around Aβ plaques, particularly around compact ones, which He found more likely to coincide with NP tau.
“The data fit very nicely with this [priming] hypothesis,” Vassar said. Vassar had described microtubule disruption in the dystrophic neurites near neuritic plaques in the 5xFAD mice, which he developed (Oakley et al., 2006). “What Lee and colleagues also realized is that tau is freed and becomes available for NP aggregate formation. That is really key,” said Vassar. Saido was also excited by this idea. “This may lead to a major paradigm shift, because neuritic dystrophies have generally been considered nonspecific accumulations of axonal proteins at pre-synapses,” he wrote.
Several researchers noted the need to gain a sharper understanding of the different aggregates’ structures. “We are examining that now,” said Lee “All we know is that they are in different parts of the neuron.” Researchers also wondered why NP tau does not form without injection of human tau seeds. Lee noted that it takes years after Aβ plaques emerge for tau aggregates to form and spread in people, and suggested that mice with Aβ plaques simply do not live long enough to develop tau pathology on their own. Alternatively, noted Patrik Brundin of the Van Andel Institute in Grand Rapids, Michigan, “there might be something specific about the three-dimensional-structure of tau from AD brains.”
Bennett said the new results complemented her own data suggesting Aβ pathology enhances formation of distinct pathological tau seeds. One intriguing difference—Bennett found increased tau seeding activity before overt Aβ deposition. “There are likely multiple mechanisms at work, possibly involving both soluble Aβ and plaques,” she wrote (see full comment below).
Bennett also noted that other factors besides tau may be present in the AD-tau extract. Lee plans to propagate AD-tau seeds in a cell-free system by incubating them with recombinant tau, or soluble tau from AD brains, to obtain a purer preparation for testing.
Lee will also extend her experiments to mouse models carrying familial AD mutations. And she hopes to use the approach to test candidate AD treatments. Brundin liked this application. “I think this is a very exciting translational model,” he said. “I suspect it will be useful for testing immunotherapies for tau, in particular, because the tau pathology is most likely very similar to that of the human brain.”—Marina Chicurel
References
News Citations
Research Models Citations
Paper Citations
- Hardy JA, Higgins GA. Alzheimer's disease: the amyloid cascade hypothesis. Science. 1992 Apr 10;256(5054):184-5. PubMed.
- Pooler AM, Polydoro M, Maury EA, Nicholls SB, Reddy SM, Wegmann S, William C, Saqran L, Cagsal-Getkin O, Pitstick R, Beier DR, Carlson GA, Spires-Jones TL, Hyman BT. Amyloid accelerates tau propagation and toxicity in a model of early Alzheimer's disease. Acta Neuropathol Commun. 2015 Mar 24;3:14. PubMed.
- Bennett RE, DeVos SL, Dujardin S, Corjuc B, Gor R, Gonzalez J, Roe AD, Frosch MP, Pitstick R, Carlson GA, Hyman BT. Enhanced Tau Aggregation in the Presence of Amyloid β. Am J Pathol. 2017 Jul;187(7):1601-1612. Epub 2017 May 10 PubMed.
- Guo JL, Narasimhan S, Changolkar L, He Z, Stieber A, Zhang B, Gathagan RJ, Iba M, McBride JD, Trojanowski JQ, Lee VM. Unique pathological tau conformers from Alzheimer's brains transmit tau pathology in nontransgenic mice. J Exp Med. 2016 Nov 14;213(12):2635-2654. Epub 2016 Oct 17 PubMed.
- Oakley H, Cole SL, Logan S, Maus E, Shao P, Craft J, Guillozet-Bongaarts A, Ohno M, Disterhoft J, Van Eldik L, Berry R, Vassar R. Intraneuronal beta-amyloid aggregates, neurodegeneration, and neuron loss in transgenic mice with five familial Alzheimer's disease mutations: potential factors in amyloid plaque formation. J Neurosci. 2006 Oct 4;26(40):10129-40. PubMed.
Further Reading
No Available Further Reading
Primary Papers
- He Z, Guo JL, McBride JD, Narasimhan S, Kim H, Changolkar L, Zhang B, Gathagan RJ, Yue C, Dengler C, Stieber A, Nitla M, Coulter DA, Abel T, Brunden KR, Trojanowski JQ, Lee VM. Amyloid-β plaques enhance Alzheimer's brain tau-seeded pathologies by facilitating neuritic plaque tau aggregation. Nat Med. 2018 Jan;24(1):29-38. Epub 2017 Dec 4 PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
RIKEN Center for Brain Science
Based on pathological chronology and genetic evidence, I and others have postulated that Aβ is like the Mafia boss and tau is the hitman. The question is, how does Aβ amyloidosis induce tauopathy? In a previous study, a team led by Brad Hyman showed that crossing tau transgenic mice with APP/PS1 transgenics accelerated tau propagation (Pooler et al., 2015), so there is indeed a connection between Aβ amyloidosis and tauopathy. What is new in this latest Nature Medicine paper by Virginia Lee’s group? They indicate that, at early seeding stages, Aβ plaques facilitate tau accumulation in the neuritic plaques rather than neurofibrillary tangle formation. This may lead to a major paradigm shift, because neuritic dystrophies generally have been considered nonspecific accumulations of axonal proteins at pre-synapses. So, the effort to understand the Aβ-tau axis has moved one step further. One remaining question is why it takes Aβ amyloidosis approximately two decades to induce tauopathy in human brain (Bateman et al., 2012). To understand how the initial tau seeds are made may be a key to answer this question.
References:
Pooler AM, Polydoro M, Maury EA, Nicholls SB, Reddy SM, Wegmann S, William C, Saqran L, Cagsal-Getkin O, Pitstick R, Beier DR, Carlson GA, Spires-Jones TL, Hyman BT. Amyloid accelerates tau propagation and toxicity in a model of early Alzheimer's disease. Acta Neuropathol Commun. 2015 Mar 24;3:14. PubMed.
Bateman RJ, Xiong C, Benzinger TL, Fagan AM, Goate A, Fox NC, Marcus DS, Cairns NJ, Xie X, Blazey TM, Holtzman DM, Santacruz A, Buckles V, Oliver A, Moulder K, Aisen PS, Ghetti B, Klunk WE, McDade E, Martins RN, Masters CL, Mayeux R, Ringman JM, Rossor MN, Schofield PR, Sperling RA, Salloway S, Morris JC. Clinical and biomarker changes in dominantly inherited Alzheimer's disease. N Engl J Med. 2012 Aug 30;367(9):795-804. PubMed.
Mass General Institute for Neurodegenerative Disease
This is an exciting paper that nicely explores how Aβ interacts to enhance tau pathology using a model system that overcomes some of the limitations of transgenic mice. In work from our lab, we recently reported that Aβ pathology interacts with tau to form more pathological tau seeds earlier on in the disease course. The findings from He et al. complement this research by demonstrating that Aβ can also spatially alter tau pathology, shifting the balance toward neuritic dystrophy tau or neurofibrillary tangle tau depending on disease stage. I think the spatio-temporal dynamic between these two hallmark pathologies is fascinating and can help explain key observations from neuropathological examination of AD cases showing Aβ in the cortical mantle precedes tau tangle formation.
I think further studies to address what forms of tau are present and how these may be differentially altered by Aβ will be exciting to look into and the results of these studies would have clear therapeutic implications. It will be interesting to determine if this neuritic-plaque-tau-to-neurofibriallary-tangle pathological progression is typical in AD or if preparing tau extracts from additional cases and brain regions will yield different results.
Make a Comment
To make a comment you must login or register.