C9ORF72 Throws a Wrench into DNA Repair Machinery
Quick Links
Hexanucleotide expansions in the C9ORF72 gene—the most common genetic cause of amyotrophic lateral sclerosis and frontotemporal dementia—mount a multipronged attack on the DNA repair system, according to a July 17 study in Nature Neuroscience. Researchers led by Mimoun Azzouz and Sherif El-Khamisy at the University of Sheffield in England reported that the repeat expansions trigger the formation of DNA-RNA hybrids, called R-loops, that break DNA. At the same time, the unusual dipeptide repeats (DPRs) translated from these expansions derail efforts to mend the damage. The researchers found evidence of broken DNA and a subpar repair response in mice expressing the expansions, and also in postmortem tissue from C9-ALS patients. They proposed that the onslaught of DNA damage in neurons ultimately leads to their demise, and that targeting the pathway could become a therapeutic strategy.
DNA damage is a common hazard inside cells, and an extensive repair system exists to lessen its toll. Neurons are acutely dependent on this repair machinery, as they cannot easily wipe the slate clean through replication (see Pan et al., 2014). Making matters worse, oxidative DNA damage increases in the brain, and repair mechanisms start to falter with age and in the context of neurodegenerative disease (Sep 2011 news; Feb 2013 conference news).
Against this backdrop, El-Khamisy and colleagues wondered if an additional stressor—C9ORF72 hexanucleotide expansions—might add fuel to the fire. These expansions of the GGGGCC sequence exist in hundreds to thousands of copies in people with ALS/FTD. Both RNA foci formed from their transcription, and the DPRs generated by their translation, reportedly inflict damage on neurons. The researchers hypothesized that due to the expansions’ repetitive nature, and the abundance of GC repeats within them, the C9ORF72 expansions could be extremely prone to folding into R-loops, a type of DNA-RNA hybrid structure that can form during transcription (Aguilera and Garcia-Muse, 2012). R-loops are known triggers of double-stranded DNA breaks (Hamperl and Cimprich, 2014).
To learn if the expansions caused R-loops, first author Callum Walker and colleagues transfected the expansions into human fetal lung fibroblasts. Then they probed with antibodies specific to R-loops and phosphorylated histone γH2AX, an established indicator of double-stranded breakages. Indeed, they found that cells expressing 102 repeats that could not be translated—and thus only formed RNA foci, not DPRs—harbored elevated numbers of R-loops and breaks. This was also the case in cells transfected with constructs that did result in the translation of 34 or 69 DPRs. Notably, overexpression of senataxin, an RNA helicase known to resolve R-loops, reduced the number of breakages and even normalized the uptick in cell death the breakages triggered. Together, the findings suggested that the repeat expansions caused R-loops, which snapped DNA and harmed cells.
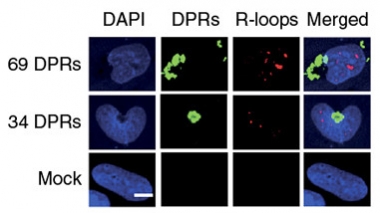
DNA, Thrown for a Loop.
Cells transfected with dipeptide repeats (green) build up R-loops (red) in their DNA. [Courtesy of Walker et al., Nature Neuroscience 2017.]
The researchers further wondered whether the repeat expansions would affect DNA repair. In both human fibroblasts and primary rat cortical neurons expressing the expansions, the researchers found the repair machinery to be profoundly hobbled. For starters, ataxia telangiectasia (ATM), the master DNA repair kinase, was hypophosphorylated and failed to activate when the researchers treated cells with DNA-damaging toxins. This led to a dismal nuclear recruitment of 53BP1, a factor that rejoins broken DNA, as well as subpar phosphorylation of another key ATM target, p53.
Through an extensive battery of biochemical and immunostaining experiments, the researchers zeroed in on the mechanisms that derailed the DNA repair machinery. The E3 ubiquitin ligase RN168 normally ubiquitylates histone H2A, an adornment that is needed to recruit 53BP1 to damaged DNA. However, in cells expressing the expansions, the researchers found RN168 tied up in p62 inclusions instead. This led to a reduction in ubiquitylated H2A and stymied 53BP1 recruitment. Interestingly, previous studies have reported that successful recruitment of 53BP1 to DNA helps sustain further ATM signaling (Lee et al., 2010). Therefore, RN168’s entrapment in p62 inclusions could potentially derail the entire DNA repair process. In support of this idea, overexpression of RN168, or depletion of p62, restored 53BP1 recruitment and reduced the number of DNA breaks in cells expressing the repeat expansions.
Strikingly, the researchers also observed R-loops, double-stranded breaks, and signs of weak ATM signaling in neurons from mice injected with viral vectors harboring the repeat expansions. These animals suffered a 20 percent loss in brainstem neurons, as well as motor deficits. The researchers proposed that DNA damage was the primary cause of this neurodegeneration, a hypothesis they will test by overexpressing senataxin and/or RN168 in the animals, El-Khamisy told Alzforum.
The researchers also found evidence of DNA in disrepair in postmortem spinal cord tissue from ALS patients, which were wrought with R-loops, double-stranded DNA breaks, and signs of ATM signaling defects.
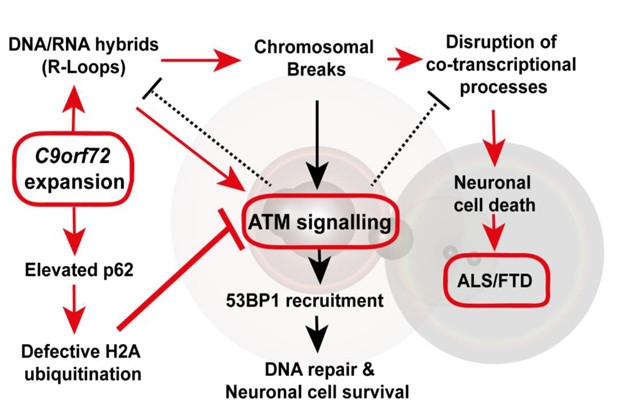
Repeat Assault. In the proposed model, C9ORF72 repeat expansions damage DNA and thwart its repair. [Courtesy of Walker et al., Nature Neuroscience 2017.]
The researchers proposed that C9ORF72 hexanucleotide expansions attacked DNA via two distinct, yet intertwined, pathways: through directly causing damage via R-loops, and by dismantling ATM-mediated DNA repair. El-Khamisy proposed that the repeat-laden RNA causes R-loops, while the DPRs manifest the p62 inclusions that sequester RN168 and disrupt repair. Interestingly, the latter pathway meshes with other recent findings implicating RN168 sequestration in p62 inclusions in the disruption of DNA repair (Wang et al., 2016).
Walker’s findings dovetail with a previous study led by Li-Huei Tsai of MIT, which reported that the ALS gene FUS is recruited to DNA breaks and helps orchestrate repair (Sep 2013 news). “The findings of the current paper … are very consistent with ours, and together make a strong argument for the role of unrepaired DNA breaks in ALS,” Tsai commented.
“This paper is particularly well done,” commented Ray Truant of McMaster University in Hamilton, Canada. “It really establishes ATM-mediated DNA repair as a common ‘node’ in neurodegenerative disease,” he said.
Truant recently reported that mutated huntingtin protein disrupted DNA repair (Maiuri et al., 2017). He added that as reactive oxidation builds in the brain with age, the efficiency of the DNA repair response could strongly influence the onset of neurodegenerative disease, a hypothesis supported by recent genome wide association studies (Bettencourt et al., 2016; Jones et al., 2017).
The study also provides researchers with a number of therapeutic targets, some of which may prove useful across neurodegenerative diseases, Truant added.—Jessica Shugart
References
News Citations
- Faulty DNA Repair Gene Leads to Cognitive Problems
- Exploring Genome Fragmentation in Neurodegeneration
- Paper Alert: FUS a Fixer of Damaged DNA
Paper Citations
- Pan L, Penney J, Tsai LH. Chromatin regulation of DNA damage repair and genome integrity in the central nervous system. J Mol Biol. 2014 Oct 9;426(20):3376-88. Epub 2014 Aug 14 PubMed.
- Aguilera A, García-Muse T. R loops: from transcription byproducts to threats to genome stability. Mol Cell. 2012 Apr 27;46(2):115-24. PubMed.
- Hamperl S, Cimprich KA. The contribution of co-transcriptional RNA:DNA hybrid structures to DNA damage and genome instability. DNA Repair (Amst). 2014 Jul;19:84-94. Epub 2014 Apr 18 PubMed.
- Lee JH, Goodarzi AA, Jeggo PA, Paull TT. 53BP1 promotes ATM activity through direct interactions with the MRN complex. EMBO J. 2010 Feb 3;29(3):574-85. Epub 2009 Dec 10 PubMed.
- Wang Y, Zhang N, Zhang L, Li R, Fu W, Ma K, Li X, Wang L, Wang J, Zhang H, Gu W, Zhu WG, Zhao Y. Autophagy Regulates Chromatin Ubiquitination in DNA Damage Response through Elimination of SQSTM1/p62. Mol Cell. 2016 Jul 7;63(1):34-48. Epub 2016 Jun 23 PubMed.
- Maiuri T, Mocle AJ, Hung CL, Xia J, van Roon-Mom WM, Truant R. Huntingtin is a scaffolding protein in the ATM oxidative DNA damage response complex. Hum Mol Genet. 2017 Jan 15;26(2):395-406. PubMed.
- Bettencourt C, Hensman-Moss D, Flower M, Wiethoff S, Brice A, Goizet C, Stevanin G, Koutsis G, Karadima G, Panas M, Yescas-Gómez P, García-Velázquez LE, Alonso-Vilatela ME, Lima M, Raposo M, Traynor B, Sweeney M, Wood N, Giunti P, SPATAX Network, Durr A, Holmans P, Houlden H, Tabrizi SJ, Jones L. DNA repair pathways underlie a common genetic mechanism modulating onset in polyglutamine diseases. Ann Neurol. 2016 Jun;79(6):983-90. Epub 2016 May 6 PubMed.
- Jones L, Houlden H, Tabrizi SJ. DNA repair in the trinucleotide repeat disorders. Lancet Neurol. 2017 Jan;16(1):88-96. PubMed.
Further Reading
News
Primary Papers
- Walker C, Herranz-Martin S, Karyka E, Liao C, Lewis K, Elsayed W, Lukashchuk V, Chiang SC, Ray S, Mulcahy PJ, Jurga M, Tsagakis I, Iannitti T, Chandran J, Coldicott I, De Vos KJ, Hassan MK, Higginbottom A, Shaw PJ, Hautbergue GM, Azzouz M, El-Khamisy SF. C9orf72 expansion disrupts ATM-mediated chromosomal break repair. Nat Neurosci. 2017 Sep;20(9):1225-1235. Epub 2017 Jul 17 PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
Picower Institute of MIT
This study is consistent with our previous work on the relationship of accumulation of DNA damage and neurodegeneration. In our paper published in 2013 in Nature Neuroscience (Wang et al.), we showed that the familial ALS gene product, FUS, a DNA/RNA binding protein, is recruited to DNA double-strand break (DSB) sites and is necessary for efficient DNA repair. Thus, the findings of the current paper that demonstrate C9ORF72 impacts DNA repair are very consistent with ours, and together make a strong argument for the role of unrepaired DNA breaks in ALS.
The one surprising finding in the current paper is that C9ORF72 repeats impair ATM activity, but increase the γH2AX signal. This is perplexing, as ATM is the key kinase that phosphorylates the histone 2A variant and creates the amplified γH2AX signal, although another member of the PI3 kinase family, DNA-PK, can also phosphorylate H2AX in post-mitotic cells such as neurons. γH2AX is known to facilitate the recruitment of DNA repair machinery to the break sites.
In our 2013 Wang et al. study, we showed that FUS loss of function caused considerable reduction in ATM activity and decreased γH2AX signal at break sites. Therefore, I am curious to know how reduced ATM activity can result in increased phosphorylation of its substrate(s), and whether DNA-PK activity compensates for the reduction in activated ATM in the context of C9ORF72 overexpression.
References:
Wang WY, Pan L, Su SC, Quinn EJ, Sasaki M, Jimenez JC, Mackenzie IR, Huang EJ, Tsai LH. Interaction of FUS and HDAC1 regulates DNA damage response and repair in neurons. Nat Neurosci. 2013 Oct;16(10):1383-91. PubMed.
University of Sheffield
I agree with Li-Huei Tsai; we were surprised too. As rightly indicated, H2AX can become phosphorylated by other nervous system kinases, such as DNA-PK, which could compensate in the absence of efficient DNA repair signalling by ATM in the context of C9ORF72 overexpression.
It should be noted that the defect we report here is in the sustained non-canonical activation of ATM signalling cascade due to inefficient H2A ubiquitylation and 53BP1 recruitment, which also correlates with reduced phosphorylation of other ATM substrates such as p53 and KAP1.
The study by Li-Huei Tsai's group back in 2013 was instrumental in setting the scene for the connection between ALS and DNA repair. Our current study reiterates this important link but also highlights important mechanistic differences by which ALS mutants impact DNA repair capacity in the nervous system.
University of Queensland
It is well established that DNA double-strand breaks and R-loops contribute to genome instability and, in turn, to pathological outcomes. What is novel about this study is that a mechanism is provided, bringing together both R-loops and DNA double-strand breaks, in explaining how a short DNA sequence, when expanded, causes two major neurodegenerative diseases, ALS and dementia.
It is also of considerable interest that this is demonstrated in both human and animal model systems. Furthermore, ATM, the protein defective in another human neurodegenerative disease, Ataxia-Telangiectasia (A-T), is impaired in its function in this system. These findings provide potential approaches to developing new therapeutic possibilities for intervention in these disorders.
Make a Comment
To make a comment you must login or register.