An Extra Strain on the Brain—α-Synuclein Seeds Tau Aggregation
Quick Links
In the brains of people with Parkinson’s disease and dementia, both tau and α-synuclein often co-aggregate. This is no case of two independent inclusions forming side-by-side, but of one seeding the other, suggests a paper in the July 3 Cell. Virginia Lee and colleagues at the University of Pennsylvania in Philadelphia report that certain forms of α-synuclein fibrils seed tau aggregation in mouse neurons in culture and in the brains of mice. However, not all kinds of α-synuclein can corrupt tau, the scientists say. They identified two different strains—one seeds α-synuclein while the other seeds tau aggregation.
Researchers who spoke with Alzforum said the findings remind them of prions. Many were not surprised to see prion-like features emerging for α-synuclein, and suspected this study might be the first of many to find multiple strains for the proteins that aggregate in Alzheimer’s, Parkinson’s, Huntington’s, amyotrophic lateral sclerosis, and related diseases. Though aggregation-prone proteins such as α-synuclein are not infectious like traditional prions, accumulating evidence suggests that within a given organism, they transfer pathogenic misfolds to neighboring, normal proteins as prions do (see ARF News story on Irwin et al., 2013; ARF News story; ARF News story; ARF News story). “This paper is a milestone paper for the field. It is very impressive, very interesting, and very exciting,” commented Mathias Jucker of the University of Tübingen, in Germany, in an email to Alzforum. “The field always expected ‘proteopathic strains’ for proteins other than the typical prions…the suggested cross-seeding mechanism is a valid hypothesis.”
Lee and first author Jing Guo had reason to suspect they could seed tau aggregation with α-synuclein fibrils. The group previously showed that α-synuclein nucleates tau aggregation in vitro (see ARF News story on Giasson et al.,), and others reported that the same occurs in human embryonic kidney cells in culture (Waxman and Giasson, 2011).
Lee’s group has already shown that transferring pathogenic α-synuclein into cells or mouse brains induces aggregation of endogenous α-synuclein (see ARF News story on Volpicelli-Daley et al., 2011; ARF News story on Luk et al., 2012. Guo attempted to seed tau aggregation in a similar fashion, transferring synthetic α-synuclein fibrils into hippocampal neurons from mouse embryos expressing mutant, aggregation-prone tau. Most of her experiments were unsuccessful, until one attempt yielded the tau inclusions she was after.
Next, Guo had to figure out why most α-synuclein preparations didn’t work, but one did. She looked to the prion field, where proteins with the same amino acid sequence often exist in multiple misfolded conformations, or “strains.” A classic way for prions to morph from one strain into another is during repeated in-vitro seeding. Researchers take a bit of the prion and use it to seed aggregation of the normal protein in a new test tube. They repeat this process several times over, thereby “passaging” a small amount of protein from one tube to the next, and sometimes the strain type changes over these passages.
Guo passaged her own synuclein fibrils, taking five or 10 percent of one prep to seed the next, in succession. Early passage fibrils had no effect on tau when incubated with the mouse hippocampal neurons, but after five to seven repeated seed-and-transfer reactions, the synuclein caused tau to aggregate. Lee and Guo labeled the early-passage fibrils strain A, and the later ones strain B. In cells, strain A typically seeded aggregation of endogenous α-synuclein. Strain B, in contrast, seeded less α-synuclein than did strain A, preferentially seeding tau aggregation. “There may be subtle differences that account for some of the conformations recruiting tau, and some not,” Lee said.
In strain B-seeded neurons, α-synuclein and tau inclusions associated closely, as if α-synuclein templated the tau aggregation (see image below). Strain A was toxic to neurons, increasing lactate dehydrogenase release and reducing metabolism in the cells within two weeks of inoculation. Neurons treated with strain B remained healthy through 18 days. Lee suspects that the larger number of aggregates in strain A cultures contributed to their demise.
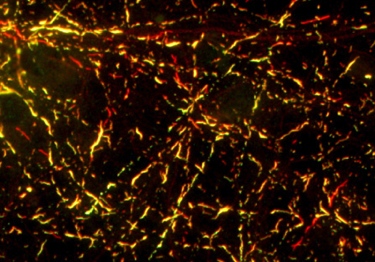
Sticking together: Mouse neurons seeded with α-synuclein strain B welled up with entwined α-synuclein (red) and tau (green) aggregates.Image courtesy of Virginia Lee, University of Pennsylvania School of Medicine, Philadelphia.
Next, Guo performed in vivo experiments, injecting each strain into the hippocampus of mice expressing mutant human tau. Normally these mice develop tau pathology around one year of age; Guo attempted to jump-start the process by injecting her strains into two- or three-month old animals. Three months after the injections, the strain A mice exhibited very few tau inclusions near the injection site. In contrast, tau aggregates appeared all over the hippocampus of mice injected with strain B. The scientists concluded that strain B α-synuclein seeded tau aggregation.
What differentiates strains A and B? Guo tried various methods to biochemically distinguish them. She discovered that they respond differently to digestion with proteinase K. Both generated five bands on a denaturing protein gel, with strain B yielding much more of the smallest fragment. That indicates strain B aggregates are less tightly bound and more easily digested, Lee said, suggesting that they are structurally different from strain A. The researchers also generated an antibody that appears to recognize strain B more strongly than strain A. That needs to be confirmed with purified preparations of the strains.
Could different a-synuclein strains exist in the human brain? In a preliminary experiment, the authors used proteinase K to digest α-synuclein from the brains of five people who had died of Parkinson’s disease dementia (PDD). Two had relatively “pure” cases of just PDD due to synucleinopathy. Three had both PDD and an Alzheimer’s diagnosis based on tau tangles and amyloid plaques. The researchers isolated detergent-soluble α-synuclein aggregates from the brains. Upon proteinase K digestion of these samples, the two “pure” PDD cases resulted in a single primary band, while the three combination cases yielded two. The results suggest to Lee and Guo that there are two variants of α-synuclein possible in the human brain, though they concede the sample size is too small for solid conclusions. Lee plans to examine more brains from cases of Parkinson’s, PDD or dementia with Lewy bodies (DLB) for further evidence of α-synuclein strains.
The possibility of there being multiple α-synuclein strains could answer a question that has long perplexed the field: why are brain pathologies and clinical symptoms so heterogeneous? For example, synucleinopathy may manifest as Parkinson’s disease, DLB, or multiple system atrophy. Perhaps the different clinical symptoms arise from slightly different strains, Lee suggested.
The prion field provides precedent for this theory, noted Claudio Soto of the University of Texas Medical School in Houston, who was not involved in the study. In the case of the original prion protein PrP, it causes different symptoms depending on which strain manifests. The same basic protein can create the ataxia and dementia of sporadic Creutzfeld-Jakob disease, the psychiatric symptoms of so-called “mad-cow” disease, or the sleeplessness of fatal familial insomnia, Soto said.
The cross-seeding also could explain mixed pathology. More than half of AD cases also exhibit α-synuclein Lewy bodies, and AD plaques and tangles often appear in the brains of people with synucleinopathies like PD or dementia with Lewy bodies (reviewed in Galpern and Lang, 2006. If certain forms of α-synuclein cross-seed tau, it could account for these mixed pathologies in some, but not all, cases.
“I thought this study was elegant and exciting,” commented Neil Cashman of the University of British Columbia in Vancouver, who was not involved in the work. However, he noted that it is very difficult to prove beyond any doubt that cross-seeding has occurred. There are always other possible explanations for why one aggregate leads to another. For example, the first type of aggregate may disrupt proteostasis or the protein degradation machinery, leading indirectly to aggregation of a second protein. Even so, the concept of cross-seeding has precedent; other researchers have suggested that some forms of amyloid can cross-seed ( Yan et al., 2007).
If researchers can learn to identify strains, they could use that information to categorize neurodegenerative diseases, the study authors suggest. For example, someday pathology might indicate a person had “synucleinopathy, strain type B” or a similar diagnosis. “This provides a reasonable approach for being able to differentiate these different syndromes on the basis of strains,” Cashman said.
Researchers who spoke with Alzforum said α-synuclein may not be the only protein that separates into strains. Amyloid-β, tau, huntingtin, SOD1, TDP-43 may also turn out to have many misfolded conformations, they suggested. In fact, researchers have already shown that tau from different human diseases causes different pathology when injected into mouse brains, suggesting the possibility of multiple strains (Clavaguera et al., 2013). Studies also suggest amyloid-β may differentiate into multiple strains (Nuallain et al., 2004). “There are going to be multiple strains in every [neurodegenerative] disorder,” Cashman predicted. “I think that will be the rule.”—Amber Dance.
References
News Citations
- In Case You Wondered: Neurodegenerative Diseases Are Not Contagious
- Proteopathic Seeds and Neurodegenerative Diseases
- Chicago—ALS Protein SOD1 Painted as Disease Template
- Do Tau "Prions" Lead the Way From Concussions to Progression?
- Tau and α-synuclein at the Nexus of Alzheimer's and Parkinson's
- Modeling Sporadic PD in a Dish?
- Toxic Synuclein Corrupts Native in Wild-Type Mice
Paper Citations
- Irwin DJ, Abrams JY, Schonberger LB, Leschek EW, Mills JL, Lee VM, Trojanowski JQ. Evaluation of potential infectivity of Alzheimer and Parkinson disease proteins in recipients of cadaver-derived human growth hormone. JAMA Neurol. 2013 Apr;70(4):462-8. PubMed.
- Giasson BI, Forman MS, Higuchi M, Golbe LI, Graves CL, Kotzbauer PT, Trojanowski JQ, Lee VM. Initiation and synergistic fibrillization of tau and alpha-synuclein. Science. 2003 Apr 25;300(5619):636-40. PubMed.
- Waxman EA, Giasson BI. Induction of intracellular tau aggregation is promoted by α-synuclein seeds and provides novel insights into the hyperphosphorylation of tau. J Neurosci. 2011 May 25;31(21):7604-18. PubMed.
- Volpicelli-Daley LA, Luk KC, Patel TP, Tanik SA, Riddle DM, Stieber A, Meaney DF, Trojanowski JQ, Lee VM. Exogenous α-synuclein fibrils induce Lewy body pathology leading to synaptic dysfunction and neuron death. Neuron. 2011 Oct 6;72(1):57-71. PubMed.
- Luk KC, Kehm V, Carroll J, Zhang B, O'Brien P, Trojanowski JQ, Lee VM. Pathological α-synuclein transmission initiates Parkinson-like neurodegeneration in nontransgenic mice. Science. 2012 Nov 16;338(6109):949-53. PubMed.
- Galpern WR, Lang AE. Interface between tauopathies and synucleinopathies: a tale of two proteins. Ann Neurol. 2006 Mar;59(3):449-58. PubMed.
- Yan J, Fu X, Ge F, Zhang B, Yao J, Zhang H, Qian J, Tomozawa H, Naiki H, Sawashita J, Mori M, Higuchi K. Cross-seeding and cross-competition in mouse apolipoprotein A-II amyloid fibrils and protein A amyloid fibrils. Am J Pathol. 2007 Jul;171(1):172-80. PubMed.
- Clavaguera F, Akatsu H, Fraser G, Crowther RA, Frank S, Hench J, Probst A, Winkler DT, Reichwald J, Staufenbiel M, Ghetti B, Goedert M, Tolnay M. Brain homogenates from human tauopathies induce tau inclusions in mouse brain. Proc Natl Acad Sci U S A. 2013 Jun 4;110(23):9535-40. PubMed.
- Halliday GM, Holton JL, Revesz T, Dickson DW. Neuropathology underlying clinical variability in patients with synucleinopathies. Acta Neuropathol. 2011 Aug;122(2):187-204. PubMed.
Further Reading
Papers
- Clavaguera F, Akatsu H, Fraser G, Crowther RA, Frank S, Hench J, Probst A, Winkler DT, Reichwald J, Staufenbiel M, Ghetti B, Goedert M, Tolnay M. Brain homogenates from human tauopathies induce tau inclusions in mouse brain. Proc Natl Acad Sci U S A. 2013 Jun 4;110(23):9535-40. PubMed.
- Halliday GM, Holton JL, Revesz T, Dickson DW. Neuropathology underlying clinical variability in patients with synucleinopathies. Acta Neuropathol. 2011 Aug;122(2):187-204. PubMed.
- Kikis EA, Gidalevitz T, Morimoto RI. Protein homeostasis in models of aging and age-related conformational disease. Adv Exp Med Biol. 2010;694:138-59. PubMed.
Primary Papers
- Guo JL, Covell DJ, Daniels JP, Iba M, Stieber A, Zhang B, Riddle DM, Kwong LK, Xu Y, Trojanowski JQ, Lee VM. Distinct α-synuclein strains differentially promote tau inclusions in neurons. Cell. 2013 Jul 3;154(1):103-17. PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
Uppsala Universitet
The concept of "strains" emanates from the prion field. It has been shown both in vitro and in animal models that the disease-associated prion protein (PrPsc) can exhibit differences in misfolding and aggregation. These different forms tend to be stable and can be propagated, again both in vitro and in vivo. The mechanism for propagation is "seeding," during which preformed aggregates with a high degree of β-sheet structure interact with normal protein and thereby transfer the folding and aggregation abnormality. The process is poorly understood. The prion strains are believed to cause different disease phenotypes. For example, variant Creutzfeldt-Jakob disease, characterized by prion variants with a typical protease K resistance pattern, are discernible from PrPsc in other prion disorders.
In this article, Guo et al. show that recombinant α-synuclein can adopt at least two fibrillar conformations, called A and B. These conformations are shown to propagate in vitro and induce different conformations in cellular systems. The most interesting and novel finding is that strain B is able to induce misfolding and aggregation of tau, the protein that makes up the neurofibrillary tangles seen in several different cerebral disorders, including Alzheimer’s and Parkinson’s diseases. This type of interaction between two proteins is called cross-seeding. Interestingly, strain B, but not strain A α-synuclein fibrils, induces tau aggregation in a transgenic mouse model by cross-seeding as well. The authors conclude there may exist many distinct fibril conformations with different abilities to cross-seed, leading to variations in forms of aggregates and that this may be a reason for differences in disease phenotypes in Parkinson's. A notable and potentially important detail is that while strain A is toxic, strain B is not. This finding shows that aggregates may be deleterious in ways other than just exerting toxicity.
The paper is interesting and the experiments very well-performed, increasing our knowledge of how proteins making up cerebral aggregates interact with each other. However, the authors ignore studies of other amyloid fibril proteins that cross-seed outside of the brain. Seeding and cross-seeding are mechanism shown with other amyloid fibril proteins, in both localized and systemic diseases, and cross-seeding has been shown to occur in animal models of systemic amyloidoses caused by amyloid A and apolipoprotein AII. In addition, and contrary to what Guo et al. state at the end of their discussion, the strain phenomenon has been proposed for peripheral amyloid diseases due to amyloid A and transthyretin. It is, therefore, very conceivable that strain formation and cross-seeding are mechanisms of general importance in many or all amyloid diseases. This is a field that deserves much more attention.
References:
O'Nuallain B, Williams AD, Westermark P, Wetzel R. Seeding specificity in amyloid growth induced by heterologous fibrils. J Biol Chem. 2004 Apr 23;279(17):17490-9. PubMed.
Lundmark K, Westermark GT, Olsén A, Westermark P. Protein fibrils in nature can enhance amyloid protein A amyloidosis in mice: Cross-seeding as a disease mechanism. Proc Natl Acad Sci U S A. 2005 Apr 26;102(17):6098-102. PubMed.
Yan J, Fu X, Ge F, Zhang B, Yao J, Zhang H, Qian J, Tomozawa H, Naiki H, Sawashita J, Mori M, Higuchi K. Cross-seeding and cross-competition in mouse apolipoprotein A-II amyloid fibrils and protein A amyloid fibrils. Am J Pathol. 2007 Jul;171(1):172-80. PubMed.
Westermark GT, Westermark P. Prion-like aggregates: infectious agents in human disease. Trends Mol Med. 2010 Nov;16(11):501-7. Epub 2010 Oct 1 PubMed.
MRC Laboratory of Molecular Biology
This work demonstrates the existence of assembled α-synuclein strains, based on the cross-seeding (or not) of tau aggregation. This is important for understanding some human neurodegenerative diseases, where α-synuclein inclusions are present in different cell types and different parts of the nervous system, sometimes in conjunction with tau inclusions. Aggregates of tau give rise to distinct human tauopathies and recent experiments have shown that strains of assembled four-repeat tau appear to exist (Clavaguera et al., 2013). Strains of assembled α-synuclein and tau provide a further link with prion diseases, where different aggregate conformations of the prion protein give rise to distinct disease phenotypes and neuropathologies.
References:
Clavaguera F, Akatsu H, Fraser G, Crowther RA, Frank S, Hench J, Probst A, Winkler DT, Reichwald J, Staufenbiel M, Ghetti B, Goedert M, Tolnay M. Brain homogenates from human tauopathies induce tau inclusions in mouse brain. Proc Natl Acad Sci U S A. 2013 Jun 4;110(23):9535-40. PubMed.
View all comments by Michel GoedertMake a Comment
To make a comment you must login or register.