Going It Alone—APP, BACE1 Take Distinct Routes to Endosome
Quick Links
Rather than tackle a criminal in your bedroom, why not prevent him from entering your home? That’s the therapeutic takeaway from a new study on intracellular trafficking of amyloid-β precursor protein (APP) and the protease that slashes it to jumpstart production of the amyloid-β peptide. Reporting online this week in the Proceedings of the National Academy of Sciences USA, researchers led by Wim Annaert of KU Leuven, Belgium, show that APP and β-secretase (BACE1) take different paths to meet in early endosomes, the primary site of Aβ generation. Whereas APP reaches these degradative compartments by clathrin-dependent mechanisms, BACE1 moves along a clathrin-independent pathway regulated by the small GTPase ADP ribosylation factor 6 (ARF6), according to the research. Though the molecular details of ARF6-mediated pathways remain murky, “What is clear from the experiments here is that the internalization of BACE1 occurs very distinctly from APP. The evidence is very convincing,” said Dudley Strickland of the University of Maryland, Baltimore. The results suggest that, rather than directly block BACE1, researchers may be able to target enzymes that smuggle it into the cell, thereby snuffing out Aβ production while leaving APP unharmed.
Whereas many labs measure Aβ production as a readout for APP processing, Annaert’s focuses on where the processing occurs—and more specifically for this study, how APP and BACE1 get from the cell surface into endosomal compartments where they meet. To get a handle on this, first author Ragna Sannerud and colleagues enhanced or blocked specific endocytic routes in HeLa cells expressing fluorescently labeled APP and BACE1 proteins, then tracked their internalization using confocal microscopy. When the researchers treated cells with epidermal growth factor (EGF) to stimulate clathrin-independent endocytosis, they saw that BACE1 got internalized faster than usual, whereas trafficking of APP and another protein, transferrin receptor, did not change. On the other hand, in cells where clathrin-dependent endocytosis was blocked using multiple strategies, uptake of APP and transferrin receptor came to a halt while BACE1 internalization proceeded normally. These data indicated that APP and BACE1 follow different routes from the cell’s surface to the endosomal compartments, where the proteins meet to do their dirty work.
In another set of experiments, Annaert’s team found a large proportion of BACE1, but not APP, co-localizing in endosomes with other membrane-bound proteins (CD59 and MHC1) that become internalized via a non-clathrin-mediated pathway. This pathway is driven by ARF6, and the researchers demonstrated that this GTPase can regulate BACE1 trafficking as well. By transfecting cells with an ARF6 mutant locked in its GTP-bound state (ARF6-Q67L)—which pushes internalized cargo into “phantom” vesicles that effectively exit the endocytic pathway (see figure)—the researchers prevented BACE1, but not APP, from reaching early endosomes.
Importantly for AD, the authors went on to show that using ARF6 mutants to manipulate BACE1 trafficking produced the expected effects on Aβ production. The ARF6-Q67L mutant slowed it. However, a GDP-locked ARF6 mutant (ARF6-T27N) that blocks recycling of proteins from endosomes to the cell surface led to the opposite outcome—more β amyloid—by forcing BACE1 and APP to hang around longer in the same compartments (see image).
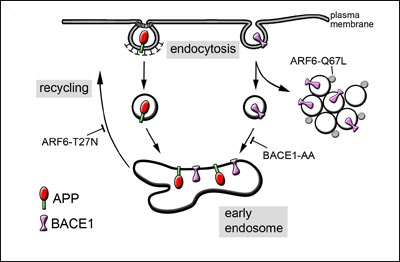
Multiple Paths to the Endosome
APP (red) and BACE1 (pink) follow distinct routes to the early endosome. Blocking specific points along the ARF6-mediated BACE internalization pathway has opposing effects on APP processing. ARF6-Q67L prevents formation of early endosomes and thus stunts Aβ production; ARF6-T27N stalls recycling of endosomes back to the cell surface, thereby increasing Aβ production. Image credit: Wim Annaert, copyright National Academy of Sciences
Finally, the authors extrapolated their findings to primary neurons with confocal and biochemical data suggesting that APP processing occurs less in the axon and much more in the somatodendritic compartment, where Aβ typically accumulates.
All told, the findings “open up the interesting idea that production of Aβ peptides is temporally and spatially regulated,” said Strickland, whose recent paper implicates a new low-density lipoprotein receptor in APP trafficking (ARF related news story on Ranganathan et al., 2011). That BACE1 and APP seem to follow distinct routes to the endosome “reveals a complexity to their cellular itinerary and raises new potential targets for pharmacological intervention,” noted Julie Donaldson of the National Heart, Lung, and Blood Institute, National Institutes of Health, Bethesda, Maryland (see full comment below). Blocking BACE1 endocytosis is probably not the way to go, Annaert suggested. Rather, “We can target the enzymes that activate or deactivate ARF6 in a way that delays trafficking of BACE1 and produces less Aβ. The aim is not to block but modulate—to keep it under control,” he said.
Giuseppina Tesco of Tufts University School of Medicine, Boston, took note of a finer point in the study—that BACE1 sorting to early endosomes required a C-terminal dileucine motif (DISLLK) known to be important for BACE1 trafficking. This domain binds Golgi-localized γ ear-containing ARF-binding (GGA) proteins, which are a focus of the Tesco lab. Her group showed that GGA3 levels are decreased in AD brain and inversely correlated with BACE1 levels (ARF related news story on Tesco et al., 2007). The current paper raises the question “of whether ARF6 regulates BACE1 endocytosis and levels via GGA3 or another adaptor,” Tesco noted in an e-mail to ARF. “It would be interesting to determine whether ARF6 levels are also changed in AD brains.” (See full comment below.) Studies of this nature are ongoing in Annaert’s lab, and preliminary data “confirm our conclusion that such endosomal pathways can be selectively affected in aging,” Annaert told ARF.—Esther Landhuis
References
News Citations
- Research Brief: Novel Sorting Protein May Affect Amyloid Production
- Stress and Aβ—The Apoptosis Connection
Paper Citations
- Ranganathan S, Noyes NC, Migliorini M, Winkles JA, Battey FD, Hyman BT, Smith E, Yepes M, Mikhailenko I, Strickland DK. LRAD3, a novel low-density lipoprotein receptor family member that modulates amyloid precursor protein trafficking. J Neurosci. 2011 Jul 27;31(30):10836-46. PubMed.
- Tesco G, Koh YH, Kang EL, Cameron AN, Das S, Sena-Esteves M, Hiltunen M, Yang SH, Zhong Z, Shen Y, Simpkins JW, Tanzi RE. Depletion of GGA3 stabilizes BACE and enhances beta-secretase activity. Neuron. 2007 Jun 7;54(5):721-37. PubMed.
Further Reading
Papers
- Ranganathan S, Noyes NC, Migliorini M, Winkles JA, Battey FD, Hyman BT, Smith E, Yepes M, Mikhailenko I, Strickland DK. LRAD3, a novel low-density lipoprotein receptor family member that modulates amyloid precursor protein trafficking. J Neurosci. 2011 Jul 27;31(30):10836-46. PubMed.
- Tesco G, Koh YH, Kang EL, Cameron AN, Das S, Sena-Esteves M, Hiltunen M, Yang SH, Zhong Z, Shen Y, Simpkins JW, Tanzi RE. Depletion of GGA3 stabilizes BACE and enhances beta-secretase activity. Neuron. 2007 Jun 7;54(5):721-37. PubMed.
- Rajendran L, Honsho M, Zahn TR, Keller P, Geiger KD, Verkade P, Simons K. Alzheimer's disease beta-amyloid peptides are released in association with exosomes. Proc Natl Acad Sci U S A. 2006 Jul 25;103(30):11172-7. PubMed.
Primary Papers
- Sannerud R, Declerck I, Peric A, Raemaekers T, Menendez G, Zhou L, Veerle B, Coen K, Munck S, De Strooper B, Schiavo G, Annaert W. ADP ribosylation factor 6 (ARF6) controls amyloid precursor protein (APP) processing by mediating the endosomal sorting of BACE1. Proc Natl Acad Sci U S A. 2011 Aug 23;108(34):E559-68. PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
Tufts University School of Medicine
This is a very interesting paper. The authors show with a variety of elegant experiments that BACE1, but not APP, endocytosis is regulated by ARF6, most likely via a clathrin-independent pathway. ARF6 is mainly found at the cell membrane and regulates endocytosis of cargo. Interestingly, ARFs bind GGAs (Golgi-localized, γ ear-containing, ADP ribosylation factor-binding proteins), which are involved in protein trafficking, and regulate GGA recruitment to the cell membrane. ARF6 seems also to regulate BACE1 levels similarly to GGA3. This work raises the questions of whether ARF6 regulates BACE1 endocytosis and levels via GGA3 or other adaptors. We have shown that GGA3 levels are decreased in AD brains and inversely correlated with BACE1 levels. It would be interesting to determine whether ARF6 levels are also changed in AD brains.
BACE1 is a primary drug target for AD therapy. However, after a decade since the discovery of β-secretase, the identification of effective BACE1 inhibitors that are active in the CNS has been very difficult. The catalytic site of BACE1 is exceptionally long, and it has been very challenging to develop small compounds that efficiently inhibit BACE1, are able to cross the blood-brain barrier, and are reasonably stable. An alternative approach to BACE1 small-molecule inhibitors is the indirect inhibition of BACE1 through the modulation of regulatory mechanisms that control BACE1 levels or BACE1 trafficking to acidic compartments. Thus, both GGA3 and ARF6 represent potential therapeutical targets to decrease Aβ production.
View all comments by Giuseppina TescoNational Heart, Lung, and Blood Institute, NIH
This study by Sannerud et al. expands our understanding of how BACE1 meets
View all comments by Julie Donaldsonup with APP in endocytic compartments to generate amyloid-β peptides.
Although much is known about clathrin-dependent forms of endocytosis and
the trafficking itinerary of plasma membrane proteins that enter cells by this
mechanism, another parallel endocytic pathway exists that handles membrane
proteins entering cells independent of the clathrin coat. There is
communication among these membrane systems and exchange of cargo, but we
are only beginning to understand the regulation and significance of these
membrane trafficking pathways in cellular function. The finding that
BACE1 enters cells through clathrin-independent endocytosis prior to
meeting up with APP in common endocytic compartments reveals a complexity
to their cellular itinerary and raises new potential targets for
pharmacological intervention. Furthermore, it highlights important
physiological functions for these “phantom” clathrin-independent endocytic
pathways that are ubiquitous but underappreciated.
University of Zurich
This elegant cell biology study adds another important piece to the puzzle of how endocytosis controls amyloid production. A few years ago, we showed that APP and BACE are internalized into early endosomes where β cleavage occurs (Rajendran et al., 2006). Here, the group of Wim Annaert shows that APP and BACE1 are internalized in distinct endocytic routes to reach the early endosome compartment. The authors show that while APP is internalized to early endosomes via a clathrin-dependent mechanism akin to that of the transferrin receptor, BACE1 is internalized via a clathrin-independent way. Certain proteins, particularly the ones associated with specialized lipid domains (lipid rafts) have been shown to be internalized via a clathrin-independent route. GPI anchored proteins and lipid binding toxins take this route to hitch their way into the interior of the cell.
While raft-dependent mechanisms have been implicated in the endocytosis of APP/ BACE1, this study is the first to show that the clathrin-independent endocytosis of BACE1 requires a GTPase called Arf6. Arf6 is a small GTPase that, similarly to Rab GTPases, regulate membrane trafficking. Interestingly, Arf6 localizes to the plasma membrane and endosomes, and through binding to β-arrestins, Arf6 regulates G protein-coupled receptor internalization and desensitization. Arf6 mediated internalization seems to regulate amyloidogenic processing of APP and Aβ production. Of course, one needs to further understand the implications of this in the light of AD. It would be interesting to check if there is an increase in Arf6-GAP (GTPase activating protein) in Alzheimer's patients. Do age-related lipid alterations (high plasma membrane cholesterol, gangliosides) increase the clathrin-independent endocytosis of BACE1? Can one therapeutically exploit this new finding by targeting specifically the Arf6 pathway?
Arf6 also controls internalization of many other cargoes, and hence, manipulating Arf6 levels or activity might not be desirable, but one could design inhibitors of BACE1 that are trafficked to this route (Rajendran et al., 2008). Cell biological investigations are starting to reveal some of the most fascinating aspects of Alzheimer's disease.
References:
Rajendran L, Honsho M, Zahn TR, Keller P, Geiger KD, Verkade P, Simons K. Alzheimer's disease beta-amyloid peptides are released in association with exosomes. Proc Natl Acad Sci U S A. 2006 Jul 25;103(30):11172-7. PubMed.
Rajendran L, Schneider A, Schlechtingen G, Weidlich S, Ries J, Braxmeier T, Schwille P, Schulz JB, Schroeder C, Simons M, Jennings G, Knölker HJ, Simons K. Efficient inhibition of the Alzheimer's disease beta-secretase by membrane targeting. Science. 2008 Apr 25;320(5875):520-3. PubMed.
View all comments by Lawrence RajendranMake a Comment
To make a comment you must login or register.