Propagation Blues? Reporter Expression Clouds Reports of Traveling Tau, Aβ
Quick Links
Updated 24 April 2015
Nefarious conformers of endogenous proteins can wander the brain cajoling their normal brethren to join the toxic fold. Right? This has emerged as a tenet of neurodegenerative disease research. However, to prove that a protein can travel from place A to place B, it is important to establish that it started off in place A. Researchers had previously relied on a handy mouse model, Nop-tTA, in which expression of a transgene was assumed to be restricted to the entorhinal cortex, to support the notion that toxic aggregates of tau and Aβ spread from there to anatomically connected areas of the brain. Now, a new study led by Joanna Jankowsky at Baylor College of Medicine in Houston reports that the Nop-tTA expression may be a good bit looser than previously realized.
As reported in the April 14 Brain Structure and Function, the researchers used a β-galactosidase reporter, and the classic blue color it generated from the sugar derivative, X-Gal, to reveal Nop-tTA expression well beyond the entorhinal cortex, including the hippocampus and many regions of the cortex. They also reported that APP expression driven by the Nop-tTA promoter was expressed in a similar pattern. The new findings do not completely nullify past studies, which had acknowledged some degree of ectopic expression, but they do make them more difficult to interpret, Jankowsky told Alzforum. Jankowsky and colleagues hope other researchers will rely on the expression atlas generated from their study when planning or interpreting future experiments using these models.
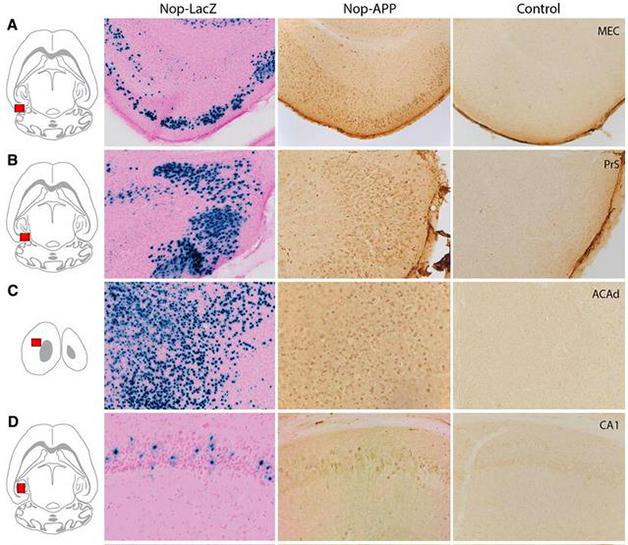
Beyond the EC. In addition to expression in the medial entorhinal cortex (A), brain sections from 6-month-old Nop-LacZ and Nop-APP mice revealed significant expression of transgenes in the presubiculum (B), dorsal anterior cingulate cortex (C), CA1 region of the hippocampus (D), and other areas.
“The study by Yetman and colleagues is nicely done, and the resolution of the images is superb,” commented Lary Walker of Emory University in Atlanta (see full comment below). “The authors convincingly show that the ‘leakage’ of transgene expression in the Nop-tTA driver line is much more extensive than was previously thought. The results undoubtedly give pause, but they don’t necessarily negate earlier conclusions that a pathogenic agent (such as Aβ or tau seeds) is being transported by neurons,” he added.
Michael Sasner of Jackson Laboratory in Bar Harbor, Maine, commented that researchers should make sure to take advantage of published expression data like Jankowsky’s when designing their experiments. “The Jankowsky group should be commended for doing this careful and important work; too often, this type of careful characterization is not done, which can lead to ambiguous or even misleading results,” he wrote (see comment below). “Ideally other journals and funding agencies will take note of this type of work and encourage some common-sense practices to promote experimental reproducibility.”
The entorhinal cortex (EC) is among the first brain regions affected by neurodegeneration in Alzheimer’s disease. Nestled between the hippocampus and other cortical regions, the EC plays a central role in memory. Restricting gene expression to the EC is an attractive strategy for understanding how its circuitry affects neighboring regions, or for measuring whether misfolded proteins such as tau and Aβ may spread from the EC outwards. In 2006, researchers led by Mark Mayford at the Scripps Research Institute in La Jolla, California, generated mice in which the tet transactivator (tTA) was expressed under control of the neuropsin (Nop) promoter, which was reportedly active primarily in the EC. When crossed with mice expressing a transgene of interest under control of a tet-responsive promoter, cells in the EC expressed the gene (see Yasuda and Mayford, 2006). Since then, several other groups have used the Nop-tTA mouse to express human APP and tau, and reported that misfolded forms of the proteins spread from the EC to the neighboring hippocampus and beyond (see Nov 2010 news; Feb 2012 news; Liu et al., 2012; and de Calignon et al., 2012). These studies acknowledged some degree of leaky expression of tau or Aβ outside of the EC, and made efforts to control for it.
With an eye toward using the Nop-tTA mice in their own studies, Jankowsky and colleagues decided to confirm the specificity of gene expression in the EC. They used the β-galactosidase gene because of its exquisite sensitivity. This reporter had been used to establish expression patterns in the original 2006 study; however, subsequent studies using different transgenes and in mice with different genetic backgrounds only looked for expression of their transgene to assess localization, Jankowsky said.
First author Michael Yetman and colleagues incubated brain slices from 2½-month-old Nop-tTA/tet-LacZ mice with X-gal to measure transgene expression throughout the brain. In agreement with the original study, the researchers found the X-gal signal was highest in the EC and neighboring pre-/parasubiculum. However, the researchers also found blue cells well outside of the EC in the hippocampus, parahippocampal regions, and dorsal isocortex, and lower levels in the visual, somatosensory, and somatomotor cortices. Green fluorescent protein (GFP) was also part of the reporter construct, and it presented a similar pattern of expression, although less pronounced than that detected by the more sensitive X-gal.
Jankowsky was surprised by these initial results, and decided to take a close accounting of transgene expression in a 7-month-old male and 7-month-old female mouse, and one 9-month-old male. The researchers found extensive expression beyond the EC. They published their results through the Rodent Brain Workbench tTA atlas. Cells in the frontal parts of the cerebral cortex had low to medium amounts of X-gal positive cells, while blue cells formed a continuous sheet across the entire parietal, temporal, and occipital regions. Within the hippocampal formation, labeled cells dotted the CA1 pyramidal layer and, to a greater degree, the subiculum. Blue cells were present in the pre- and parasubiculum, as well as in the perihinal areas and of course the EC, which had the most pronounced expression in the medial zone. Blue cells were also found in the olfactory bulb, amygdala, and striatum, while other regions, including the substantia nigra and much of the midbrain and brainstem, were relatively spared.
Why did Jankowsky and colleagues find more extensive reporter expression than researchers did in the original 2006 study? While it is impossible to know for sure, Jankowsky said the major difference between the two studies was not the technique used to detect the reporter, but the genetic background of the animals. In Mayford’s 2006 study, the Nop-tTA mice were on a hybrid C57BL/6/DBA background that was backcrossed one time to C57BL/6. By the time Jankowsky used them for the current study, they had been backcrossed at least six times to C57BL/6. Jankowsky said differences in genetic background have been known to influence the expression patterns of transgenes, and this may explain the differences in their data.
The researchers next crossed the Nop-tTA mice to mice expressing a tet-responsive human APP gene with two familial AD mutations, and measured APP expression in brain slices using the 6E10 antibody. They found that for the most part, APP expression mimicked what they had observed with X-gal: Regions with the highest density of blue cells also had the highest APP expression, and vice versa (see image above).
The researchers hypothesized that low levels of APP expression in regions such as CA1 could ultimately give rise to amyloid plaques, but that it would take longer for plaques to form in these regions than in areas with more pronounced APP expression, such as the EC. Indeed, the researchers found that amyloid plaques (as measured by silver staining) were restricted to the EC and pre-/parasubiculum in 9-month-old, double-transgenic mice, but by 15 months of age they were also present in the CA1 and dentate gyrus regions of the hippocampus, as well as the visual, auditory, and retrosplenial cortex. All of these regions had displayed sparse LacZ expression in the β-gal reporter animals and moderate or low APP expression in younger Nop-tTA/APP mice. However, all these regions connect to the entorhinal circuit, making it difficult to distinguish whether the Aβ came from the EC or was produced by local expression of APP.
By 21 months, amyloid deposits were detected throughout the neocortex and hippocampal formation. Plaques were largely absent from regions that showed low or no X-gal labeling and no APP expression, such as CA3, striatum, and insular cortex—notable exceptions included the lateral septal nucleus and orbital cortex, which produced no APP but did have dense plaques. These regions had well-known axonal connections with other areas of the brain, including but not limited to the EC, that did express APP. The researchers concluded that in those areas, Aβ was likely delivered by afferent axons and released into local synapses. The same might occur in regions that did express APP. How much local expression of APP versus axonal transmission contributed to the appearance of amyloid pathology in the model was hard to assess, Jankowsky said.
Do these findings throw a wrench into previous studies based on this model? Not necessarily, Jankowsky said. In 2010, researchers led by Lennart Mucke at the University of California, San Francisco, used the Nop-tTA model to express mutant human APP. Using the same 6E10 antibody that Jankowsky used to stain brain sections, Mucke and colleagues found transgenic APP expression predominantly inside, but also outside of the EC. In 13-month-old mice, the researchers observed amyloid plaques in the EC and the dentate gyrus— the latter being an area where both Jankowsky and Mucke had observed only very sparse APP expression. Mucke concluded that at least in the dentate gyrus, amyloid deposits likely derived from Aβ released from presynaptic terminals of neurons projecting from the EC.
Two studies used the Nop-tTA to track the spread of tau in the rodent brain (see Feb 2012 news). Researchers led by Karen Duff at Columbia University in New York crossed the Nop-tTA line to Tg4510 mice, which express a tet-responsive human tau gene harboring the P301L mutation. The offspring were a hybrid of C57BL/6 and FVB genetic backgrounds. In that study, the researchers found that misfolded and hyperphosphorylated tau appeared in the EC at 10 months and was then found in the dentate gyrus and CA1 regions of the hippocampus by 22 months. To check whether tau was expressed in the dentate gyrus, Duff and colleagues used laser capture microdissection to isolate cells from that region and checked for human tau mRNA. They did find a low level of expression there, but concluded that it was insufficient to account for the amount of tau protein in the region. They concluded that tau was likely transported to the dentate gyrus through neurons in the perforant pathway, which projects from the EC.
A paper from Bradley Hyman’s lab at Massachusetts General Hospital in Charlestown used the same mice and came to similar conclusions. Hyman's team scrutinized tau expression beyond the EC using a combination of fluorescence in situ hybridization (FISH) and immunofluorescence to simultaneously measure human tau mRNA and human tau protein at the single cell level. They found that in the dentate gyrus, CA1, CA3, and anterior cingulate cortex, many of the cells harboring tau protein did not express human tau mRNA, although some did. They concluded that a significant fraction of misfolded tau protein found in these regions traveled there from the EC.
A subsequent paper from Mucke’s lab reported that while pathological forms of tau appeared in the dentate gyrus of these mice as they aged, low levels of non-pathological forms of human tau were expressed at all ages in the double-transgenic mice as well as in mice expressing tetO-htau, without the Nop-tTA driver. This suggested a leakiness from the tet-responsive promoter (see Harris et al., 2012), although whether this low level of expression would have eventually produced tau pathology is unknown.
Jankowsky said that the major conclusion drawn from Duff’s and Hyman’s studies—that misfolded tau can propagate via neuronal circuitry from one brain region to another—is likely correct, and has been supported by other studies using different methods. “I don’t think that expression outside of the EC accounts for everything, but because the system has these potential caveats, I think it’s hard to disambiguate what is real spread from what is just slower accumulation by cells with less transgenic protein,” she said.
Duff agreed that leaky expression is certainly a problem in this mouse model, but added that its low levels were unlikely to account for the large amounts of aggregated tau researchers found outside of the EC in their studies. A careful accounting of tau expression levels was crucial to draw these conclusions, Duff said, which is why her and Hyman’s studies focused only on regions connected directly to the entorhinal circuit. “We could make a statement about tau propagation in that one region, but we should be very careful if we want to extrapolate beyond that area,” she said. Hyman told Alzforum that the possibility of ectopic expression is an issue researchers face when relying on animal models such as Nop-tTA. "Performing extensive control experiments in these cases is crucial, and those undertaken in our study were sufficient to support the conclusions," he said.
David Borchelt of the University of Florida in Gainesville commented that the study should compel researchers to more carefully assess expression patterns in their mouse models. “As pointed out by Yetman and colleagues, it is quite possible that the strain background of the mice modifies expression patterns for this tTA transgene. Thus, it is difficult to know whether the publications that used these mice to conclude that Aβ and tau spread trans-synaptically are flawed,” he wrote (see full comment below). “Still, the Yetman study really sets a standard for how one goes about defining the pattern of transgene expression.”—Jessica Shugart
References
News Citations
- Insidious Spread of Aβ: More Support for Synaptic Transmission
- Mice Tell Tale of Tau Transmission, Alzheimer’s Progression
Paper Citations
- Yasuda M, Mayford MR. CaMKII activation in the entorhinal cortex disrupts previously encoded spatial memory. Neuron. 2006 Apr 20;50(2):309-18. PubMed.
- Liu L, Drouet V, Wu JW, Witter MP, Small SA, Clelland C, Duff K. Trans-synaptic spread of tau pathology in vivo. PLoS One. 2012;7(2):e31302. PubMed.
- de Calignon A, Polydoro M, Suárez-Calvet M, William C, Adamowicz DH, Kopeikina KJ, Pitstick R, Sahara N, Ashe KH, Carlson GA, Spires-Jones TL, Hyman BT. Propagation of tau pathology in a model of early Alzheimer's disease. Neuron. 2012 Feb 23;73(4):685-97. PubMed.
- Harris JA, Koyama A, Maeda S, Ho K, Devidze N, Dubal DB, Yu GQ, Masliah E, Mucke L. Human P301L-mutant tau expression in mouse entorhinal-hippocampal network causes tau aggregation and presynaptic pathology but no cognitive deficits. PLoS One. 2012;7(9):e45881. PubMed.
External Citations
Further Reading
No Available Further Reading
Primary Papers
- Yetman MJ, Lillehaug S, Bjaalie JG, Leergaard TB, Jankowsky JL. Transgene expression in the Nop-tTA driver line is not inherently restricted to the entorhinal cortex. Brain Struct Funct. 2015 Apr 14; PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
University of Florida
I think the data nicely show that is possible for expression of tTA in the Nop-tTA mice to extend beyond what has been reported for these animals. As pointed out by Yetman et al., it is quite possible that the strain background of the mice modifies expression patterns for this tTA transgene. Thus, it is difficult to know whether the publications that used these mice to conclude that Aβ and tau spread trans-synaptically are flawed. Still, the Yetman study really sets a standard for how one goes about defining the pattern of transgene expression, and the data cast a considerable shadow on the published Aβ/tau work. It now becomes incumbent on the authors of these studies to more carefully prove an absence of APP or tau transgene expression in the regions of the brain that were said to have acquired pathology by trans-synaptic spreading.
View all comments by David R. BorcheltEmory University
The study by Yetman and colleagues is nicely done, and the resolution of the images is superb. The authors convincingly show that the “leakage” of transgene expression in the Nop-tTA driver line is much more extensive than was previously thought. The results undoubtedly give pause, but they don’t necessarily negate earlier conclusions that a pathogenic agent (such as Aβ or tau seeds) is being transported by neurons (indeed, the authors concur with Harris et al., 2010, that Aβ appears to be transported to regions distant from the primary sites of transgene expression); whether specific agents are conveyed trans-synaptically to connected neurons in vivo remains to be determined. The conclusion “that leak rather than (or in addition to) trans-synaptic spread could have contributed” to the anatomical distribution of lesions in the AD mouse models seems reasonable in light of present knowledge. Numerous capable researchers are addressing the problem of cellular trafficking of seeds in vitro; I am especially looking forward to studies that nail down the mechanism of lesion spread within the living brain.
References:
Harris JA, Devidze N, Verret L, Ho K, Halabisky B, Thwin MT, Kim D, Hamto P, Lo I, Yu GQ, Palop JJ, Masliah E, Mucke L. Transsynaptic progression of amyloid-β-induced neuronal dysfunction within the entorhinal-hippocampal network. Neuron. 2010 Nov 4;68(3):428-41. PubMed.
View all comments by Lary WalkerThe Jackson Lab
The Jankowsky group should be commended for doing this careful and important work; too often, this type of careful characterization is not done, which can lead to ambiguous or even misleading results.
Anyone selecting a mouse for a specific application should take advantage of the existing information resources. In addition to those mentioned in the paper, the Mouse Genome Informatics creportal presents annotated expression data on more than 2,200 distinct cre-expressing strains, representing more than 600 different genetic transgenic drivers and more than 400 knock-in drivers. The Allen Brain Atlas data portal presents brain serial sections of in situ analysis of cre reporters for more than 100 different cre lines, all of which are available from a mouse repository. We at JAX are also developing data on off-target expression in the developing embryo and in adult organs other than the brain (all information is available in the creportal).
While tet-expressing strains have not had the same level of analysis, there is a similar Klk8-tTA strain (same construct, different founder line from the Mayford lab) to that used in this paper with expression analysis presented in the Allen Brain Atlas data portal. This mouse is available from the MMRRC repository.
Ideally, other journals and funding agencies will take note of this type of work and encourage some common-sense practices to promote experimental reproducibility. It is very inefficient for each lab to have to do this type of analysis, so having central resources such as the Allen Brain Atlas and the Mouse Genome Informatics data are essential. The use of congenic, or at least standard, genetic backgrounds should be encouraged. Preferably, each project would assay multiple transgenic lines, to minimize the chance that the result is an artifact of aberrant expression in a single founder line. Where possible, use a mouse from a repository or donate your mouse to a repository, so that other labs can easily obtain the mouse and reproduce/extend your work. The simplest and most effective change would be to require that publications unambiguously identify the strain(s) that were used, ideally with a unique ID; very often it is not clear from a publication what experimental tools were used. PubMed has the “LinkOut” feature that can provide a quick link to the resources (e.g. mouse models). These practices don’t have to be costly, but can make results significantly more meaningful.
View all comments by Michael SasnerYetman et al. report a straightforward experiment crossing Mayford’s Nop line S (which they obtained five years ago) with a reporter line. The initial line was selected from multiple lines because of its relative restriction to neurons of interest, predominantly but not exclusively in the medial entorhinal cortex. Yetman et al. show data from a handful of animals – using semiquantitative scoring – to illustrate the extent to which the promoter drives a reporter mostly but not exclusively in the entorhinal cortex. We (and I imagine all groups that have used this line) are in broad agreement with this observation, although in our hands the extent to which the reporter is expressed is not as widespread as they report. Still, our line has also been separately bred from the parent strain for ~five years, and some drift could easily occur.
Where we differ is the implication that the idea of tau propagation, put forward by our lab and by Karen Duff’s lab in 2012 (de Calignon et al., 2012; Liu et al., 2012) based in part on results from this line, should be questioned based on Yetman et al.’s observations that some neurons outside of the entorhinal cortex express the promoter. The critical observation by our lab and Duff’s is in the dentate gyrus, the neurons that receive projections from the entorhinal cortex become immunopositive for human tau protein as the mice age. We did not simply assume that entorhinal cortex was the only place that expressed human tau in the transgene. We did extensive control experiments to conclusively demonstrate that the dentate gyrus cells that contained human tau protein did not express human tau mRNA.
Five lines of evidence support this critical point, using a variety of techniques, have been presented in DeCalignon et al. and subsequently (Polydoro et al., 2014):
1. A fluorescent protein reporter line demonstrates absence of expression of a reporter protein in the dentate gyrus.
2. Double in-situ hybridization/immunofluorescence demonstrates absence of human tau mRNA in the exact neurons that are immunopositive for human tau protein
3. Laser capture microdissection of neurons that contain human tau mRNA versus those that did not, with or without being immunopositive for human tau protein, showed by a sensitive qPCR specific for human tau mRNA that none could be detected in the neurons that were in situ negative, tau protein positive.
4. The number of neurons that become immunopositive for tau increases with age and corresponds to the pace at which the dentate gyrus terminal zone degenerates. This observation would be difficult to explain if it were due to mistaken expression. These experiments directly ask if tau mRNA is expressed in the relevant cells, and the answer is unequivocally no.
5. To this list we can now add a fifth line of evidence: Yetman et al.’s table 1 shows a score of zero for the dentate gyrus, suggesting that with yet another and potentially more sensitive reporter, there is no misexpression in dentate gyrus.
It is worth noting that tau propagation has been seen in vitro and in vivo by multiple labs using multiple techniques, including direct injection of human tau protein and viral-based human tau expression (Dujardin et al., 2014; Wu et al., 2013).
Yetman et al. make an interesting point with regard to the possibility that strain or colony drift have changed the properties of the animals they examine. In our hands there are scattered neurons in other fields, although by far the predominant expression is in the entorhinal cortex and adjacent pre- and parasubiculum. In other transgenic lines we have observed phenotype drift over time and breeding, and we remain committed to carefully documenting expression patterns in the exact animals we use for experiments on an ongoing basis. Yetman et al. make the excellent point that this is important in any experimental paradigm where anatomical patterns of expression matter, and we definitely agree.
References:
Yetman MJ, Lillehaug S, Bjaalie JG, Leergaard TB, Jankowsky JL. Transgene expression in the Nop-tTA driver line is not inherently restricted to the entorhinal cortex. Brain Struct Funct. 2015 Apr 14; PubMed.
de Calignon A, Polydoro M, Suárez-Calvet M, William C, Adamowicz DH, Kopeikina KJ, Pitstick R, Sahara N, Ashe KH, Carlson GA, Spires-Jones TL, Hyman BT. Propagation of tau pathology in a model of early Alzheimer's disease. Neuron. 2012 Feb 23;73(4):685-97. PubMed.
Liu L, Drouet V, Wu JW, Witter MP, Small SA, Clelland C, Duff K. Trans-synaptic spread of tau pathology in vivo. PLoS One. 2012;7(2):e31302. PubMed.
Polydoro M, Dzhala VI, Pooler AM, Nicholls SB, McKinney AP, Sanchez L, Pitstick R, Carlson GA, Staley KJ, Spires-Jones TL, Hyman BT. Soluble pathological tau in the entorhinal cortex leads to presynaptic deficits in an early Alzheimer's disease model. Acta Neuropathol. 2014 Feb;127(2):257-70. Epub 2013 Nov 24 PubMed.
Dujardin S, Lécolle K, Caillierez R, Bégard S, Zommer N, Lachaud C, Carrier S, Dufour N, Aurégan G, Winderickx J, Hantraye P, Déglon N, Colin M, Buée L. Neuron-to-neuron wild-type Tau protein transfer through a trans-synaptic mechanism: relevance to sporadic tauopathies. Acta Neuropathol Commun. 2014 Jan 30;2(1):14. PubMed.
Wu JW, Herman M, Liu L, Simoes S, Acker CM, Figueroa H, Steinberg JI, Margittai M, Kayed R, Zurzolo C, Di Paolo G, Duff KE. Small Misfolded Tau Species Are Internalized via Bulk Endocytosis and Anterogradely and Retrogradely Transported in Neurons. J Biol Chem. 2013 Jan 18;288(3):1856-70. PubMed.
Allen Institute for Brain Science
Gladstone Institutes/UCSF
The Gladstone Institutes
The paper by Yetman et al. provides a valuable resource for investigators planning experiments with this neuropsin-tTA driver line, and the availability of this data in an open-access resource should be lauded. While bringing important attention to the fact that this neuropsin-tTA mouse line is not perfect in its restriction to the EC, the findings of Yetman et al. should not be seen in any way as “casting a shadow” on the results or interpretation of our APP/Aβ and hTau studies using the Nop-tTA line (Harris et al., 2010; Harris et al., 2012).
In fact, we systematically assessed expression of APP and hTau protein, and APP mRNA, in both the neuropsin-tTA/TetO-APP or TetO-hTau double-transgenic mice as well as in the singly transgenic TetO-APP and TetO-hTau across the cortex and hippocampus. We used this information to interpret our results with care (see, e.g., Figure 1 and Supplemental Figures 2-4 in Harris et al., 2010). Our analysis revealed essentially similar results as in the current paper. Thus, we very purposefully referred to expression in this mouse line as “predominantly” in the EC, a decision recognized by Yetman and co-authors.
The paper by Yetman et al. does bring to light a very important caveat on the use of transgenic mouse lines. The point should be made that not only do driver lines need to be carefully analyzed, as was done nicely in this paper, but reporter lines must be as well, as they carry their own intrinsic expression patterns (TetO lines can be leaky). We found this to be an issue with both the TetO-APP and TetO-hTau lines; there was true leaky expression of APP in the absence of tTA in CA1 of the TetO-APP line and of hTau in the dentate gyrus granule cells of the TetO-hTau mice. Leak of hTau in TetO-hTau singly transgenic mice was age-dependent. At young ages, some singly transgenic mice did not express any hTau, but at older ages, all mice did (Harris et al., 2012). This leakiness could be avoided by using the second generation tetO promoter (TRE-tight) in our hands (unpublished).
However, in agreement with our paper, Yetman et al. confirmed that the Nop-tTA/TetO-APP mice do not show detectable expression of APP in granule cells of the dentate gyrus, providing at least one pathway with which to test the transmission hypothesis of Aβ from point A (entorhinal cortex) to point B (granule cells of the dentate gyrus). In fact, the authors of this study agreed with our conclusion that the spread of Aβ pathology from the EC to the molecular layer of the DG likely arose through release from perforant path terminals. We also found alterations in calbindin and fos expression within DG granule cells and physiological changes specifically at EC to DG synapses which suggest a network-based transmission of pathology (here meaning molecular and functional changes) from one region to another without a clear understanding yet as to the exact mechanism of this transfer.
Unlike with the APP mice, we did detect hTau in DG granule cells of singly transgenic TetO-hTau mice. In light of this result, our opinion, as also stated in Harris et al., 2012, was that it was impossible to definitively conclude that hTau was transferred from the EC to the DG. We did, however, observe pathological forms of Tau in these same cells only in the Nop-tTA/TetO-hTau animals.
Thus, the main findings and conclusions of our APP/Aβ and hTau papers remain unchanged in light of this new paper: APP overexpression predominantly in the EC-hippocampal network is sufficient to drive age-dependent molecular and functional pathologies and cognitive impairments similar to those seen with brain-wide overexpression of APP/Aβ. On the other hand, although overexpression of hTau in a similar spatial pattern causes pathologies within this network, it is not sufficient to induce age-dependent cognitive decline, at least up to 16 months of age.
References:
Harris JA, Koyama A, Maeda S, Ho K, Devidze N, Dubal DB, Yu GQ, Masliah E, Mucke L. Human P301L-mutant tau expression in mouse entorhinal-hippocampal network causes tau aggregation and presynaptic pathology but no cognitive deficits. PLoS One. 2012;7(9):e45881. PubMed.
Make a Comment
To make a comment you must login or register.