A Secretase Complex Built for Two
Quick Links
The α- and γ-secretases sequentially cut the amyloid precursor protein. Scientists used to think these enzymes worked autonomously in the plasma membrane, but new research published in the December 21 Journal of Cell Biology suggests they work together as a team. Using biochemical and immunohistochemical techniques, scientists led by Dennis Selkoe, Brigham and Women’s Hospital, Boston, report that the two enzymes hook up to form an active multi-protease complex. They also report that γ-secretase forms similar but separate complexes with β-secretase to churn out Aβ.
The results shed new light on the basic biology of these secretases. “We think these findings add a new level of elegance and complexity to this type of proteolysis,” Selkoe told Alzforum. “The results suggest that over eons a more efficient mechanism of cleavage has evolved.” This could be a broader phenomenon, as other membrane proteases—for example Site-2-protease and signal peptide peptidase—could exist in similar complexes, added first author Allen Chen, also at the Brigham.
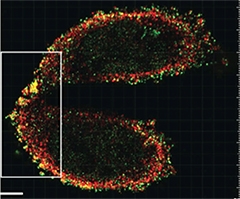
Dynamic Duo:
Super-resolution microscopy reveals that γ-secretase (red) couples with α-secretase (green) in the plasma membrane of CHO cells. [©Chen et al., 2015. Originally published in J Cell Biol.]
The data counter the view that γ-secretase cleaves many different substrates in an unregulated way, wrote Bart De Strooper, KU Leuven in Belgium (see full comment below).
Enzymes that process transmembrane proteins such as APP and Notch, which is important for cell fate determination, do so in an ordered fashion. First, either α- or β-secretase release the ectodomain of the full-length protein in a process known as shedding, then γ-secretase cleaves the intramembrane portion of the C-terminal fragment. However, Chen previously found that γ-secretase also bound to full-length APP (Chen et al., 2010). He wondered if it might do so while α-secretase was still attached.
To test this, Chen and colleagues checked whether the proteins formed complexes in lysates from Chinese hamster ovary (CHO) cells. Isolating α-secretase by immunoprecipitation also pulled down γ-secretase, and vice versa. Likewise, co-IP pulled down both proteins in neuroblastoma cell lines and in tissues from mouse brains. By labeling surface proteins with biotin and pulling down with streptavidin, the researchers found that the secretases interact in the plasma membrane, where both proteases are known to process substrates. Size exclusion chromatography confirmed the association, indicating that the two secretases exist in the same high-molecular weight complex. To test whether these complexes were proteolytically active, the researchers immunoprecipitated either ADAM10 (aka α-secretase) or γ-secretase and looked for enzymatic activity of the other. In both cases they found it, suggesting the two secretases are functional in the complex.
Chen and colleagues then tested if the two proteins co-localized at the plasma membrane. Super-resolution microscopy, which resolves objects as close as 100 nm, revealed secretase pairs dotting the periphery of CHO cells (see image above). The secretases also co-localized in the hippocampi of three-month-old wild-type mice together with the postsynaptic marker Homer. This suggested that the two secretases may meet up at synapses.
Given that the data pointed to a multiprotease complex, the researchers wondered if inhibiting γ-secretase would influence ADAM10 activity. Adding a γ-secretase inhibitor to CHO or human neuroblastoma cells caused a dose-dependent uptick in sAPPα, the soluble ectodomain fragment released by ADAM10, along with a corresponding drop in sAPPβ generated by β-secretase.
Curiously, γ-secretase inhibition also led to more APP and more BACE1 at the plasma membrane. The former might explain the elevated sAPPα, while the latter might reduce sAPPβ production, since β-secretase functions best in intracellular acidic vesicles. As for why these proteins gather at the cell membrane upon blocking γ-secretase, Chen speculated that when cells detect reduction in proteolytic products such as the Notch intracellular domain or the APP equivalent AICD, they route more substrate to the membrane for processing. Modulators of γ-secretase, which alter the substrate cleavage site but have no effect on overall enzymatic activity or Notch processing, had no effect on sAPPα.
The researchers next wondered what glues the multi-secretase complex together. Several tetraspanins—proteins that corral membrane components—have been found to associate separately with α- and γ-secretases (Xu et al., 2009; Haining et al., 2012; Wakabayashi et al., 2009). RNAi knock–down of tetraspanin 12 and 17 in CHO cells lessened the interaction between ADAM10 and γ-secretase. It also quieted the secretase cross-talk, meaning a γ-secretase inhibitor less effectively upped sAPPα and reduced sAPPβ production. “That knocking down two tetraspanins … causes less complex formation and decreases the γ—α-secretase feedback strongly suggests that proper complex formation is necessary for the feedback to work properly,” Selkoe wrote to Alzforum.
What about BACE1, which along with γ-secretase releases Aβ from APP? In tissue from mouse brains, γ-secretase and BACE1 co-immunoprecipitated, suggesting that they, too, pair up in the membrane. However, since ADAM10 and BACE1 appeared separately, the researchers believe the β–/γ-complexes are independent from those formed by α- and γ-secretase.
Together the data shore up the basic biological knowledge of these enzymes. “This study contains a huge number of biochemical and microscopy data supporting a close interaction between the γ−secretase complex and α-secretase and β-secretase, respectively,” said Paul Saftig, Christian-Albrechts-University in Kiel, Germany. The work raises questions about where the components of these complexes are formed in human neurons, and how they are transported, he said.
Do these data have therapeutic implications? De Strooper noted that pharmaceutical companies and funders of AD research are wary about potential side effects of globally inhibiting these activities. “A dynamic clustering of proteases in the membrane might provide specificity and regulation to γ-secretase processing,” he said. “Targeting these complexes specifically is an important aim for drug discovery in the next decade.”—Gwyneth Dickey Zakaib
References
Paper Citations
- Chen AC, Guo LY, Ostaszewski BL, Selkoe DJ, LaVoie MJ. Aph-1 associates directly with full-length and C-terminal fragments of gamma-secretase substrates. J Biol Chem. 2010 Apr 9;285(15):11378-91. PubMed.
- Xu D, Sharma C, Hemler ME. Tetraspanin12 regulates ADAM10-dependent cleavage of amyloid precursor protein. FASEB J. 2009 Nov;23(11):3674-81. PubMed.
- Haining EJ, Yang J, Bailey RL, Khan K, Collier R, Tsai S, Watson SP, Frampton J, Garcia P, Tomlinson MG. The TspanC8 Subgroup of Tetraspanins Interacts with A Disintegrin and Metalloprotease 10 (ADAM10) and Regulates Its Maturation and Cell Surface Expression. J Biol Chem. 2012 Nov 16;287(47):39753-65. PubMed.
- Wakabayashi T, Craessaerts K, Bammens L, Bentahir M, Borgions F, Herdewijn P, Staes A, Timmerman E, Vandekerckhove J, Rubinstein E, Boucheix C, Gevaert K, De Strooper B. Analysis of the gamma-secretase interactome and validation of its association with tetraspanin-enriched microdomains. Nat Cell Biol. 2009 Nov;11(11):1340-6. PubMed.
Further Reading
Primary Papers
- Chen AC, Kim S, Shepardson N, Patel S, Hong S, Selkoe DJ. Physical and functional interaction between the α- and γ-secretases: A new model of regulated intramembrane proteolysis. J Cell Biol. 2015 Dec 21;211(6):1157-76. PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
University Kiel
This study certainly contains a huge amount of biochemical and microscopy data supporting a close interaction between the γ-secretase complex and α-secretase and between γ- and β-secretases. As the authors point out, several γ-secretase interactome studies did not identify ADAM10 (aka α-secretase), suggesting an indirect association (this is also true the other way around … for α-secretase interactome studies). The authors explain that tetraspanins, which are known to be essential for ADAM10 and γ-secretase activity, are possible mediators of these interactions. I would assume that these super secretase complexes are part of the tetraspanin web and that this secretase-secretase interaction helps determine the assembly of the super complex, its half-life, its activity, and its substrate handling.
There are—as always—numerous questions raised by this study, including how cleavage is regulated in such webs, what are the components of these complexes in human neurons, when are the complexes formed, and how they are intracellularly transported. Is such an assembly of a membrane protease complex and substrates also relevant for other proteases? The role of the γ-secretase complexes could also be analyzed in genetic models, and it would be nice to clarify their function(s) in a more physiological context.
UK Dementia Research Institute@UCL and VIB@KuLeuven
The paper of Chen and co-authors is highly interesting, supporting the concept that γ-secretases interact with α- and β-secretases and that the enzymes are clustered in specific subdomains in the cellular membranes. It remains to be sorted out to what extent the different γ-secretase complexes behave similarly, how many different combinations of proteases and, why not, of specific substrates, are possible in these membrane microdomains. A dynamic clustering of proteases and substrates like APP, Notch, N-Cadherin in multimolecular complexes in the plane of the membrane might actually provide specificity and regulation to γ-secretase processing (De Strooper and Annaert, 2010); contradicting the dogmatic view in the field that γ-secretases cleave by default many different substrates in an unregulated way.
This view has frightened the pharmaceutical industry (and funders of AD research) because of the potential side effects when globally inhibiting these activities. However, the work here, together with other work showing differential expression and activity of the different complexes and regulation of their activity by membrane compartmentalization (see Vetrivel et al., 2005; Thathiah et al., 2013), shows that much still has to be learned.
Tetraspanins, as suggested in this work but also before, might play a central role in the regulation of γ-secretase substrate specificity by compartmentalization of the enzymes in membrane microdomains (Wakabayashi et al., 2009; Dunn et al., 2010). Understanding this intriguing compartmentalization in the context of the cell will teach us that these enzymes have very precise functions. Targeting these complexes specifically is an important aim for drug discovery in the next decade.
References:
De Strooper B, Annaert W. Novel research horizons for presenilins and γ-secretases in cell biology and disease. Annu Rev Cell Dev Biol. 2010 Nov 10;26:235-60. PubMed.
Vetrivel KS, Cheng H, Kim SH, Chen Y, Barnes NY, Parent AT, Sisodia SS, Thinakaran G. Spatial segregation of gamma-secretase and substrates in distinct membrane domains. J Biol Chem. 2005 Jul 8;280(27):25892-900. PubMed.
Thathiah A, Horré K, Snellinx A, Vandewyer E, Huang Y, Ciesielska M, De Kloe G, Munck S, De Strooper B. β-arrestin 2 regulates Aβ generation and γ-secretase activity in Alzheimer's disease. Nat Med. 2013 Jan;19(1):43-9. PubMed.
Wakabayashi T, Craessaerts K, Bammens L, Bentahir M, Borgions F, Herdewijn P, Staes A, Timmerman E, Vandekerckhove J, Rubinstein E, Boucheix C, Gevaert K, De Strooper B. Analysis of the gamma-secretase interactome and validation of its association with tetraspanin-enriched microdomains. Nat Cell Biol. 2009 Nov;11(11):1340-6. PubMed.
Dunn CD, Sulis ML, Ferrando AA, Greenwald I. A conserved tetraspanin subfamily promotes Notch signaling in Caenorhabditis elegans and in human cells. Proc Natl Acad Sci U S A. 2010 Mar 30;107(13):5907-12. Epub 2010 Mar 10 PubMed.
Make a Comment
To make a comment you must login or register.