12 Phosphorylation-Mimicking Mutations Twist Tau into AD Filaments
Quick Links
The infamous paired helical filaments (PHFs) of tau that sprawl across the brain in Alzheimer’s disease are the end products of a multiyear process. What drives tau to form these specific filaments in the first place, and how is hyperphosphorylation involved? As described in a reviewed preprint posted August 26 in eLife, researchers led by Michel Goedert and Sjors Scheres of the MRC Laboratory of Molecular Biology, Cambridge, U.K., get closer to some answers. By mutating select residues of tau into aspartate to mimic phosphorylation, the scientists zeroed in on a combination of 12 residues that, when mutated, goad tau into pairing up into helical filaments in vitro. At their core, these PHFs folded into the same back-to-back C-shaped structures found in the Alzheimer’s brain. All 12 of the mutated residues in this “PAD12” tau lie outside of the fibril core, within the so-called fuzzy coat. In vitro, PAD12-tau monomers morphed into full-fledged AD PHFs when exposed to tau filaments extracted from AD brain, and the resulting fibrils could then be used as seeds to further propagate AD-tau filaments both in vitro and in tau biosensor cell lines.
- PAD12, a recombinant form of tau with 12 phosphomimetic mutations, can be coaxed to fold into AD-tau fibrils in vitro.
- These PAD12 fibrils seed tau fibrillization in vitro and in cell lines.
- All 12 mutations lie in the fibril fuzzy coat.
“[The authors] illustrate for the first time that a full-length recombinant tau construct is capable of taking on the PHF fold found in AD,” commented Calina Glynn of the Rosalind Franklin Institute in Didcot, U.K. “These mutations to the fuzzy coat—a region not often considered when pursuing structures of tau filaments—appear to be critical to forming the PHF structure from full-length tau.”
Many groups have tried to generate recombinant tau fibrils in vitro to match those found in the brains of people with AD and other neurodegenerative diseases. These attempts have all fallen short in some way. Tau fibrillized in vitro with the help of negatively charged molecules such as RNA or heparin differ in structure from that found in the brain, as do filaments seeded in cell lines (Abskharon et al., 2022; Zhang et al., 2019; Tarutani et al., 2023). For their part, Goedert and Scheres previously reported that tau297-391—a version of tau truncated at both its N- and C-terminal ends—could form AD-like PHFs (Dec 2023 news). Addition of regions N- or C-terminal, which ultimately make up its fuzzy coat, to the ordered core of tau fibrils hampered fibril formation in vitro. They found that tau containing the carboxy end could form protofilaments if that C-terminus contained four phosphomimetic mutations. Alas, these protofilaments balked at partnering with others of their kind to form PHFs (Lövestam et al., 2022).
For their new study, first author Sofia Lövestam and colleagues set their sights on full-length tau, investigating whether adding phosphomimetic mutations to tau’s N-terminal fuzzy coat might coax the entire protein to fold into PHFs. They targeted tau’s proline-rich region, which contains residues that are notoriously phosphorylated in people with AD. To mimic this phosphorylation, they mutated another eight amino acids to aspartate—T181, S202, T205, T212, S214, T217, T231, and S235—in addition to their original C-terminal quartet—S396, S400, T403, and S404. With this, “Phosphomimetics of AD,” aka PAD12 tau, was born (image below). After seven days under the just-right conditions— intermittent shaking in a buffer of potassium phosphate and potassium citrate—the majority of PAD12 had folded into PHFs which, under the cryo-electron microscope, appeared to have the same structure as AD-tau. This was true whether PAD12 was made entirely of 3R-tau, or of both 3R- and 4R-tau isoforms. However, when PAD12 comprised only 4R-tau, it produced single protofilaments with a core structure resembling the chronic traumatic encephalopathy (CTE) fold.
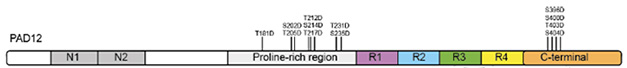
Fuzzy Phosphomimetics. The recombinant PAD12 protein includes eight phosphomimetic mutations in tau’s proline-rich region, and four in its C-terminus. All 12 mutations lie in the fuzzy coat of tau fibrils, outside of the core. [Courtesy of Lövestam et al., eLife, 2025.]
PAD12-tau PHFs looked like AD-tau PHFs, but did they behave like them? To test this, Lövestam first used brain-derived AD-tau filaments to seed PAD12 monomers. With a nudge from the AD-tau fibrils, PAD12 morphed into AD-tau PHFs within hours. What’s more, these PAD12 fibrils then acted as potent seeds themselves, quickly templating PAD12 monomers to fold into the AD-tau configuration (image below). This seeding property is distinct from that of wild-type tau: According to a study led by Mei Hong at MIT in Cambridge, it reportedly can be seeded into AD-tau PHFs with copious amounts of AD brain-derived filaments, but is incapable of acting as a seed itself in further rounds of aggregation (Duan et al., 2024).
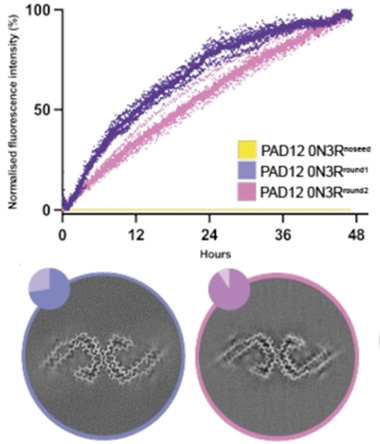
Potent PAD12. Without a fibril seed, PAD12 monomers did not fibrillize within 48 hours (yellow, top). Exposure to brain-derived AD filaments provoked rapid fibrillization (purple). The resulting PAD12 fibrils then seeded more PAD12 monomers to assemble into fibrils (pink). Both rounds of seeded aggregation produced PHFs with the AD core fold, as seen via cryo-EM (bottom). [Courtesy of Lövestam et al., eLife, 2025.]
PAD12-tau fibrils also potently seeded tau in biosensor cell lines, where they provoked HA-tagged tau297-331 monomers to aggregate with higher potency than did tau297-331 fibrils made in vitro.
Why does this phosphomimetic work so well? The authors think that hyperphosphorylation of specific residues in tau’s fuzzy coat may reduce the energy needed to nucleate new filaments. “In addition, the negative charges of hyperphosphorylated tau in filaments may actively recruit soluble tau monomers with little phosphorylation through electrostatic attraction,” they wrote. This could explain Hong’s previous observation that the assembly of wild-type full-length tau with large amounts of AD brain-derived seeds only worked for a single round of seeding, they wrote. The resulting filaments, which were not hyperphosphorylated, were then unable to seed more wild-type tau.
The findings complement those of a recent study led by Marc Diamond at the University of Texas Southwestern Medical Center in Dallas, which used alanine scanning across the tau protein to pinpoint residues that were critical for incorporation of tau into growing fibrils in different neurodegenerative tauopathies (Jan 2025 news). Though the fibrils generated in these cell-culture-based experiments may not have been a spot-on match with those in the brain, the technique accurately distinguished folds taken in different tauopathies, and singled out residues involved with the formation of each unique fold, including some that wind up in the fuzzy coats of tau fibrils.
Glynn noted that the ability of PAD12 tau to fold into a bona fide, fuzzy-coat-wearing AD PHF will allow labs without access to brain-derived material to develop small molecules and fibril-targeted therapeutics. “The true test of impact here will be on the ability of other groups to reproduce these findings, particularly without the need for cryo-EM structure confirmation for every batch of fibrils generated,” she wrote.
David Eisenberg and Michael Sawaya of University of California, Los Angeles, agreed. “If PAD12 is found to consistently form PHFs, this construct is bound to find many applications in understanding tau action and for drug development,” they wrote.—Jessica Shugart
References
News Citations
- Transient Filament Yields AD and CTE Tau Fibrils
- Cell-Based Assay Matches Tau Strains to Neuropathological Diagnosis
Paper Citations
- Abskharon R, Sawaya MR, Boyer DR, Cao Q, Nguyen BA, Cascio D, Eisenberg DS. Cryo-EM structure of RNA-induced tau fibrils reveals a small C-terminal core that may nucleate fibril formation. Proc Natl Acad Sci U S A. 2022 Apr 12;119(15):e2119952119. Epub 2022 Apr 4 PubMed.
- Zhang W, Falcon B, Murzin AG, Fan J, Crowther RA, Goedert M, Scheres SH. Heparin-induced tau filaments are polymorphic and differ from those in Alzheimer's and Pick's diseases. Elife. 2019 Feb 5;8 PubMed.
- Tarutani A, Lövestam S, Zhang X, Kotecha A, Robinson AC, Mann DM, Saito Y, Murayama S, Tomita T, Goedert M, Scheres SH, Hasegawa M. Cryo-EM structures of tau filaments from SH-SY5Y cells seeded with brain extracts from cases of Alzheimer's disease and corticobasal degeneration. FEBS Open Bio. 2023 Aug;13(8):1394-1404. Epub 2023 Jul 7 PubMed.
- Lövestam S, Koh FA, van Knippenberg B, Kotecha A, Murzin AG, Goedert M, Scheres SH. Assembly of recombinant tau into filaments identical to those of Alzheimer's disease and chronic traumatic encephalopathy. Elife. 2022 Mar 4;11 PubMed.
- Duan P, Dregni AJ, Xu H, Changolkar L, Lee VM, Lee EB, Hong M. Alzheimer's disease seeded tau forms paired helical filaments yet lacks seeding potential. J Biol Chem. 2024 Sep;300(9):107730. Epub 2024 Aug 28 PubMed.
Further Reading
No Available Further Reading
Primary Papers
- Lövestam S, Wagstaff JL, Katsinelos T, Shi J, Freund SM, Goedert M, Scheres SH. Twelve phosphomimetic mutations induce the assembly of recombinant full-length human tau into paired helical filaments. 2025 Jul 23 10.1101/2024.10.23.619846 (version 2) bioRxiv.
Annotate
To make an annotation you must Login or Register.
Comments
Rosalind Franklin Institute
Lövestam et al. illustrate, for the first time, that a full-length recombinant tau construct is capable of taking on the PHF fold found in AD. The protein construct, which harbors 12 phosphomimetic mutations, more closely resembles the charges associated with the phosphorylation pattern in tau filaments isolated from AD brain. These mutations to the fuzzy coat—a region not often considered when pursuing structures of tau filaments—appear to be critical to forming the PHF structure from full-length tau. The authors hypothesize that these mutations precede filament formation by altering the conformation of monomeric tau, which is supported by both the PAD12 construct and by phosphorylated tau taking on a different electrophoretic mobility to that of non-hyperphosphorylated tau, first illustrated by Lindwall and Cole (Lindwall and Cole, 1984).
The fuzzy coat has long been suspected to play a role in seed-competence, which is supported here with seeded material also bearing seed-competence. This opens the possibility for seeds to be tracked in cells, tissues, and animal models and to be distinguished from newly formed aggregates. The fuzzy coat also appears to limit the possibility of larger assemblies of three to four protofilaments forming, which had been observed with variability for the tau297-391 construct (Lövestam et al., 2022; Glynn et al., 2024). These larger assemblies may influence specific labelling and suffer from quaternary-structure-dependent variability in fibril properties.
From previous work with tau297-391, after the work from Lövestam et al., we found that not only did shaking speed and vessel structure influence tau filament structure, but the instrument used was also critical. As noted by the authors here, as well as ourselves, physical factors such as mechanism of shaking and heating can be challenging to control across instruments and lead to wildly different structures. In this work, the more readily available Eppendorf ThermoMixer C is a welcome change as an economical choice because it is already used by labs and is more affordable.
This work has the potential to spur further AD research with new cell lines, animal models, filament binders, and mechanistic experiments that were previously inaccessible without large quantities of pure material. The ability to produce the pure PHF fold with an intact fuzzy coat would extend the ability to create small molecule binders and specific labels beyond researchers with access to AD material to anyone who has access to readily available biochemistry equipment. The true test will be on the ability of other groups to reproduce these findings, particularly without the need for cryo-EM structure confirmation for every batch of fibrils generated, which will be necessary initially to validate the robustness of the protocol.
References:
Lindwall G, Cole RD. The purification of tau protein and the occurrence of two phosphorylation states of tau in brain. J Biol Chem. 1984 Oct 10;259(19):12241-5. PubMed.
Lövestam S, Koh FA, van Knippenberg B, Kotecha A, Murzin AG, Goedert M, Scheres SH. Assembly of recombinant tau into filaments identical to those of Alzheimer's disease and chronic traumatic encephalopathy. Elife. 2022 Mar 4;11 PubMed.
Glynn C, Chun JE, Donahue CC, Nadler MJ, Fan Z, Hyman BT. Reconstitution of the Alzheimer's Disease Tau Core Structure from Recombinant Tau297-391 Yields Variable Quaternary Structures as Seen by Negative Stain and Cryo-EM. Biochemistry. 2024 Jan 16;63(2):194-201. Epub 2023 Dec 28 PubMed.
UCLA
UCLA-DOE Molecular Biology Institute
Structures of hyperphosphorylated tau fibrils extracted from postmortem brains of Alzheimer’s disease patients invariably show a double J-shaped structured core formed by the microtubule binding region (“paired helical filaments,” PHFs), surrounded by a less-structured fuzzy coat. In contrast, recombinant tau protein constructs form a bewildering variety of fibril polymorphs, documented in detail by previous work by Lövestam and colleagues (Lövestam et al., 2022) and illustrated in the Amyloid Atlas. Now Lövestam et al. report that a recombinant phosphomimetic mutant sequence of tau robustly forms the Alzheimer’s PHF polymorph fibril.
This mutant protein, termed PAD12, replaces a total of 12 serine and threonine residues in tau’s fuzzy coat region with aspartate. The PHFs formed by PAD12 seed tau biosensor cells. The authors suggest that their findings imply hyperphosphorylation of tau may be sufficient for the formation of the tau fold. They found that replacement of the same residues by glutamate also leads to PHFs.
Using NMR, the authors argue that glutamate replacements interfere with the formation of an intramolecular interface between two IVYK motifs, one being part of the VQIVYK steric zipper structure (Sawaya et al., 2007) of the first amyloid intermediate (Lövestam et al., 2024). That is, preventing formation of an intramolecular interface enables formation of a nucleating intermolecular interface. This suggestion is consistent with the free energy landscape of amyloid polymorphs being extremely rough; thus small changes may eliminate regions of roughness, in this case smoothing the path toward PHF formation.
If PAD12 is found to consistently form PHFs, this construct is bound to find many applications both for understanding tau action and for drug development.
References:
Lövestam S, Koh FA, van Knippenberg B, Kotecha A, Murzin AG, Goedert M, Scheres SH. Assembly of recombinant tau into filaments identical to those of Alzheimer's disease and chronic traumatic encephalopathy. Elife. 2022 Mar 4;11 PubMed.
Sawaya MR, Sambashivan S, Nelson R, Ivanova MI, Sievers SA, Apostol MI, Thompson MJ, Balbirnie M, Wiltzius JJ, McFarlane HT, Madsen AØ, Riekel C, Eisenberg D. Atomic structures of amyloid cross-beta spines reveal varied steric zippers. Nature. 2007 May 24;447(7143):453-7. PubMed.
Lövestam S, Li D, Wagstaff JL, Kotecha A, Kimanius D, McLaughlin SH, Murzin AG, Freund SM, Goedert M, Scheres SH. Disease-specific tau filaments assemble via polymorphic intermediates. Nature. 2024 Jan;625(7993):119-125. Epub 2023 Nov 29 PubMed.
OSU College of Medicine
This study refines the author’s approach for synthesizing paired helical filaments from recombinant tau protein. The innovation is to mimic tau phosphorylation, a major post-translational modification of human tau protein that attains stoichiometries of 6 to 9 moles phosphate per mole tau protein in Alzheimer’s disease. In addition to its biological effects (e.g., weakening tau affinity for microtubules), phosphorylation lowers tau solubility by neutralizing its normally positive net charge at physiological pH. Much of what we know about phosphorylation effects come from “pseudophosphorylation” models—the insertion of negatively charged Asp or Glu residues in place of hydroxyamino acids at known phosphorylation sites. For example, we found that site-specific pseudophosphorylation did not trigger tau aggregation on its own, but stabilized filaments once they formed, thereby lowering the concentration of tau required to support aggregation (Necula and Kuret, 2004).
Lövestam et al. extended this technique by incorporating 12 pseudophosphorylation sites into recombinant tau to create PAD12. In the context of 0N4R tau substrate, PAD12 lowered net charge from +12 to zero, thereby maximizing the solubility-lowering effect of this approach. Nonetheless, as in previous pseudophosphorylation studies, lowering net charge was insufficient to trigger tau aggregation by itself over tractable time scales. Moreover, neither amino acid sequence (Chiti and Dobson, 2017) nor pseudophosphorylation alone (Mammeri et al., 2024) provides the information needed for tau to adopt the specific folding patterns that characterize different diseases.
However, by combining pseudophosphorylation with their previously established environmental conditions of vigorous agitation in the presence of specific salt formulations (Lövestam et al., 2022), the authors were able to trigger aggregation while influencing folding to produce PHF conformer in high yield and purity. Under these conditions, the solubility-lowering effects of pseudophosphorylation provide important advances in performance.
First, it is now possible to drive aggregation of full-length tau into its PHF conformer. The resulting filaments contain the “fuzzy coat,” and so are less prone to clumping, thereby improving storage life and performance in morphological assays. Not only is the fuzzy coat the principal site of phosphorylation, it also mediates interaction with various cellular components, including certain canonical chaperones (Mok et al., 2018). It is now possible to interrogate these aspects of biology in the context of PHF morphology.
Second, the authors demonstrate improved performance in seeding assays, which now can be propagated through multiple cycles at lower bulk tau concentrations. This has implications for seeding the conformers of other tauopathies currently available in much lower quantities than PHFs.
Third, the system can be used to test other post-translational modifications in the context of PHF formation. For example, although the stoichiometry of lysine modifications appears to be low (e.g., dephosphorylation of brain-derived PHFs restores migration patterns on gel electrophoresis nearly identical to modification-free recombinant tau isoforms, Goedert et al., 1992), they have been predicted to affect polymorphism through steric hindrance (Arakhamia et al., 2020).
Finally, despite being conducted under nonphysiological conditions and tau concentrations, the authors show how specific interactions among nucleation motifs may influence aggregation pathway. Overall, the new methods provide important new tools for advancing study of tau lesion formation in tauopathies.
References:
Arakhamia T, Lee CE, Carlomagno Y, Duong DM, Kundinger SR, Wang K, Williams D, DeTure M, Dickson DW, Cook CN, Seyfried NT, Petrucelli L, Fitzpatrick AW. Posttranslational Modifications Mediate the Structural Diversity of Tauopathy Strains. Cell. 2020 Feb 20;180(4):633-644.e12. Epub 2020 Feb 6 PubMed.
Chiti F, Dobson CM. Protein Misfolding, Amyloid Formation, and Human Disease: A Summary of Progress Over the Last Decade. Annu Rev Biochem. 2017 Jun 20;86:27-68. Epub 2017 May 12 PubMed.
Goedert M, Spillantini MG, Cairns NJ, Crowther RA. Tau proteins of Alzheimer paired helical filaments: abnormal phosphorylation of all six brain isoforms. Neuron. 1992 Jan;8(1):159-68. PubMed.
Lövestam S, Koh FA, van Knippenberg B, Kotecha A, Murzin AG, Goedert M, Scheres SH. Assembly of recombinant tau into filaments identical to those of Alzheimer's disease and chronic traumatic encephalopathy. Elife. 2022 Mar 4;11 PubMed.
Mammeri NE, Dregni AJ, Duan P, Hong M. Structures of AT8 and PHF1 phosphomimetic tau: Insights into the posttranslational modification code of tau aggregation. Proc Natl Acad Sci U S A. 2024 Mar 5;121(10):e2316175121. Epub 2024 Feb 26 PubMed.
Mok SA, Condello C, Freilich R, Gillies A, Arhar T, Oroz J, Kadavath H, Julien O, Assimon VA, Rauch JN, Dunyak BM, Lee J, Tsai FT, Wilson MR, Zweckstetter M, Dickey CA, Gestwicki JE. Mapping interactions with the chaperone network reveals factors that protect against tau aggregation. Nat Struct Mol Biol. 2018 May;25(5):384-393. Epub 2018 Apr 30 PubMed.
Necula M, Kuret J. Pseudophosphorylation and glycation of tau protein enhance but do not trigger fibrillization in vitro. J Biol Chem. 2004 Nov 26;279(48):49694-703. PubMed.
Make a Comment
To make a comment you must login or register.