ACE Supercharges Microglia to Shrink Plaques
Quick Links
Mouse microglia got a genetic tune-up. On 9 June in Nature Aging, researchers led by Warren G. Tourtellotte at Cedars-Sinai Medical Center in Los Angeles reported that expressing human angiotensin-converting enzyme (ACE) in mouse microglia boosts their uptake of Aβ, clearing out plaques in a model of Alzheimer’s disease. These souped-up microglia helped preserve vulnerable neurons and put the brakes on memory decline.
- Microglia expressing ACE take up more Aβ.
- They shrink plaques in 5xFAD mice.
- This slows learning and memory losses.
Though best known for cleaving peptides that regulate blood pressure, ACE also has a history in Alzheimer’s research. It modulates immune responses in myeloid cells (Cao et al., 2020). Genome-wide association and proteogenomic analysis have flagged this enzyme as a risk gene for Alzheimer’s. Rare coding variants in ACE have also been linked to AD and accelerate Aβ-fueled neurodegeneration in mice (Wightman et al., 2021; Bellenguez et al., 2022; Dec 2024 news, Oct 2020 news). ACE has also been shown to degrade Aβ when overexpressed in cell culture (Hemming and Selkoe, 2005).
Because other AD risk genes, such as APOE and TREM2, can affect how microglia engulf and degrade Aβ, first author Andrew Gomez and colleagues wondered whether ACE might do something similar—and, in so doing, abate amyloid pathology.
Alas, microglia don’t express much ACE. “We have examined ACE in mouse microglia at baseline and in microglia activated by Aβ and have seen no detectable levels of expression,” Tourtellotte told Alzforum. He is looking into expression in human brain. “ACE is upregulated in the macrophages present in granulomas, suggesting that ACE induction may be part of an endogenous response to boost their immune function. It makes sense that this might also occur in human microglia as well,” he wrote to Alzforum.
How might that affect the cells? To test this, the researchers needed a way to switch on the human ACE gene in these cells. They engineered mice to carry a silent copy of the gene, kept in check by an upstream STOP cassette flanked by LoxP sites—short DNA sequences recognized by the Cre enzyme. They crossed these mice with another strain that expresses Cre in myeloid cells upon tamoxifen treatment. Once the mice got tamoxifen in their chow, Cre excised the STOP cassette, flipping on ACE expression in all myeloid cells, including microglia.
After yet another cross, with 5xFAD mice that build up widespread amyloid deposition, the scientists turned on the transgene. Over time, cell turnover ensured that endogenous peripheral immune cells, sans ACE, soon replaced those carrying the transgene, while the longer-lived microglia of the brain retained ACE expression. In other words, by 9 months of age, only ACE-expressing microglia remained and they swarmed around plaques in greater numbers than their unmodified counterparts in control mice—and they swallowed up twice as much Aβ. Inside these microglia, amyloid was shuttled into the CD68-positive endolysosomes. In ACE microglia, these swelled up and contained nearly twice as much amyloid (image below).
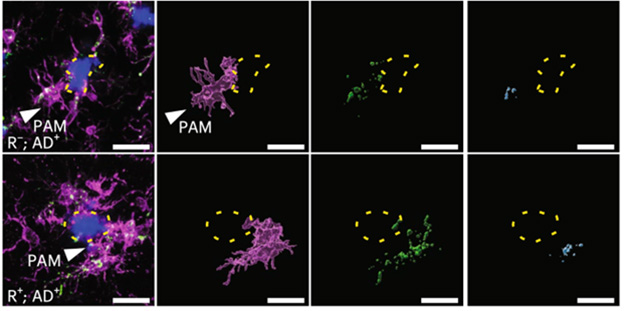
ACE Augmentation. Microglia (purple) cluster around amyloid plaques (blue). ACE microglia (bottom), engulf nearly twice as much amyloid (blue, right panel), and traffic it to CD68-positive endolysosomes (green) for degradation. [Courtesy of Gomez et al., Nature Aging, 2025.]
Wolfing down all that extra amyloid paid off. 5xFAD mice with ACE microglia had fewer amyloid deposits throughout the brain. The burden was reduced in the CA1 region of the hippocampus, the striatum, the parietal association cortex, and the amygdala (image below). Without the boost from ACE, amyloid wreaked havoc on neurons in these regions, triggering degeneration, whereas ACE-enhanced microglia helped preserve these areas.

ACE in the Brain. Mice with ACE-expressing microglia (right) had fewer amyloid deposits (white) in the parietal association cortex (PtA), CA1, and amygdala (AMG), compared to those without ACE (left). [Courtesy of Gomez et al., Nature Aging, 2025.]
The benefits extended to behavior. In the Barnes maze, a memory test where mice learn to locate a hidden escape hatch, 1-year-old mice with ACE-microglia found the exit faster during a five-day training period. Two days later, when tested on recall, they still found the exit quickly.
A key question, Tourtellotte noted, is whether expressing ACE in mouse microglia boosts a natural immune mechanism or activates a pathway that these cells wouldn’t normally use. “Either way, it seems ACE expression in mouse microglia can ‘supercharge’ their immune function,” he wrote. To what extent, if at all, the findings in this artificial overexpression system apply to the aging human brain is unclear.
Curiously, prior research has suggested that ACE inhibitors—widely used antihypertensives—may reduce brain amyloid burden, and epidemiological studies hint that some, particularly those that cross the blood–brain barrier, may modestly lower dementia risk, though results have been mixed and the mechanism unclear (Scotti et al., 2021; Sink et al., 2009).—George R. Heaton
George Heaton is a freelance writer in Durham, North Carolina.
References
News Citations
- ‘Proteogenomics’ Ties Genes That Control 38 Proteins to Alzheimer’s
- New ACE Variant Speeds Neurodegeneration in Alzheimer’s Mice
Research Models Citations
Paper Citations
- Cao DY, Saito S, Veiras LC, Okwan-Duodu D, Bernstein EA, Giani JF, Bernstein KE, Khan Z. Role of angiotensin-converting enzyme in myeloid cell immune responses. Cell Mol Biol Lett. 2020;25:31. Epub 2020 May 25 PubMed.
- Wightman DP, Jansen IE, Savage JE, Shadrin AA, Bahrami S, Holland D, Rongve A, Børte S, Winsvold BS, Drange OK, Martinsen AE, Skogholt AH, Willer C, Bråthen G, Bosnes I, Nielsen JB, Fritsche LG, Thomas LF, Pedersen LM, Gabrielsen ME, Johnsen MB, Meisingset TW, Zhou W, Proitsi P, Hodges A, Dobson R, Velayudhan L, 23andMe Research Team, Sealock JM, Davis LK, Pedersen NL, Reynolds CA, Karlsson IK, Magnusson S, Stefansson H, Thordardottir S, Jonsson PV, Snaedal J, Zettergren A, Skoog I, Kern S, Waern M, Zetterberg H, Blennow K, Stordal E, Hveem K, Zwart JA, Athanasiu L, Selnes P, Saltvedt I, Sando SB, Ulstein I, Djurovic S, Fladby T, Aarsland D, Selbæk G, Ripke S, Stefansson K, Andreassen OA, Posthuma D. A genome-wide association study with 1,126,563 individuals identifies new risk loci for Alzheimer's disease. Nat Genet. 2021 Sep;53(9):1276-1282. Epub 2021 Sep 7 PubMed. Correction.
- Bellenguez C, Küçükali F, Jansen IE, Kleineidam L, Moreno-Grau S, Amin N, Naj AC, Campos-Martin R, Grenier-Boley B, Andrade V, Holmans PA, Boland A, Damotte V, van der Lee SJ, Costa MR, Kuulasmaa T, Yang Q, de Rojas I, Bis JC, Yaqub A, Prokic I, Chapuis J, Ahmad S, Giedraitis V, Aarsland D, Garcia-Gonzalez P, Abdelnour C, Alarcón-Martín E, Alcolea D, Alegret M, Alvarez I, Álvarez V, Armstrong NJ, Tsolaki A, Antúnez C, Appollonio I, Arcaro M, Archetti S, Pastor AA, Arosio B, Athanasiu L, Bailly H, Banaj N, Baquero M, Barral S, Beiser A, Pastor AB, Below JE, Benchek P, Benussi L, Berr C, Besse C, Bessi V, Binetti G, Bizarro A, Blesa R, Boada M, Boerwinkle E, Borroni B, Boschi S, Bossù P, Bråthen G, Bressler J, Bresner C, Brodaty H, Brookes KJ, Brusco LI, Buiza-Rueda D, Bûrger K, Burholt V, Bush WS, Calero M, Cantwell LB, Chene G, Chung J, Cuccaro ML, Carracedo Á, Cecchetti R, Cervera-Carles L, Charbonnier C, Chen HH, Chillotti C, Ciccone S, Claassen JA, Clark C, Conti E, Corma-Gómez A, Costantini E, Custodero C, Daian D, Dalmasso MC, Daniele A, Dardiotis E, Dartigues JF, de Deyn PP, de Paiva Lopes K, de Witte LD, Debette S, Deckert J, Del Ser T, Denning N, DeStefano A, Dichgans M, Diehl-Schmid J, Diez-Fairen M, Rossi PD, Djurovic S, Duron E, Düzel E, Dufouil C, Eiriksdottir G, Engelborghs S, Escott-Price V, Espinosa A, Ewers M, Faber KM, Fabrizio T, Nielsen SF, Fardo DW, Farotti L, Fenoglio C, Fernández-Fuertes M, Ferrari R, Ferreira CB, Ferri E, Fin B, Fischer P, Fladby T, Fließbach K, Fongang B, Fornage M, Fortea J, Foroud TM, Fostinelli S, Fox NC, Franco-Macías E, Bullido MJ, Frank-García A, Froelich L, Fulton-Howard B, Galimberti D, García-Alberca JM, García-González P, Garcia-Madrona S, Garcia-Ribas G, Ghidoni R, Giegling I, Giorgio G, Goate AM, Goldhardt O, Gomez-Fonseca D, González-Pérez A, Graff C, Grande G, Green E, Grimmer T, Grünblatt E, Grunin M, Gudnason V, Guetta-Baranes T, Haapasalo A, Hadjigeorgiou G, Haines JL, Hamilton-Nelson KL, Hampel H, Hanon O, Hardy J, Hartmann AM, Hausner L, Harwood J, Heilmann-Heimbach S, Helisalmi S, Heneka MT, Hernández I, Herrmann MJ, Hoffmann P, Holmes C, Holstege H, Vilas RH, Hulsman M, Humphrey J, Biessels GJ, Jian X, Johansson C, Jun GR, Kastumata Y, Kauwe J, Kehoe PG, Kilander L, Ståhlbom AK, Kivipelto M, Koivisto A, Kornhuber J, Kosmidis MH, Kukull WA, Kuksa PP, Kunkle BW, Kuzma AB, Lage C, Laukka EJ, Launer L, Lauria A, Lee CY, Lehtisalo J, Lerch O, Lleó A, Longstreth W Jr, Lopez O, de Munain AL, Love S, Löwemark M, Luckcuck L, Lunetta KL, Ma Y, Macías J, MacLeod CA, Maier W, Mangialasche F, Spallazzi M, Marquié M, Marshall R, Martin ER, Montes AM, Rodríguez CM, Masullo C, Mayeux R, Mead S, Mecocci P, Medina M, Meggy A, Mehrabian S, Mendoza S, Menéndez-González M, Mir P, Moebus S, Mol M, Molina-Porcel L, Montrreal L, Morelli L, Moreno F, Morgan K, Mosley T, Nöthen MM, Muchnik C, Mukherjee S, Nacmias B, Ngandu T, Nicolas G, Nordestgaard BG, Olaso R, Orellana A, Orsini M, Ortega G, Padovani A, Paolo C, Papenberg G, Parnetti L, Pasquier F, Pastor P, Peloso G, Pérez-Cordón A, Pérez-Tur J, Pericard P, Peters O, Pijnenburg YA, Pineda JA, Piñol-Ripoll G, Pisanu C, Polak T, Popp J, Posthuma D, Priller J, Puerta R, Quenez O, Quintela I, Thomassen JQ, Rábano A, Rainero I, Rajabli F, Ramakers I, Real LM, Reinders MJ, Reitz C, Reyes-Dumeyer D, Ridge P, Riedel-Heller S, Riederer P, Roberto N, Rodriguez-Rodriguez E, Rongve A, Allende IR, Rosende-Roca M, Royo JL, Rubino E, Rujescu D, Sáez ME, Sakka P, Saltvedt I, Sanabria Á, Sánchez-Arjona MB, Sanchez-Garcia F, Juan PS, Sánchez-Valle R, Sando SB, Sarnowski C, Satizabal CL, Scamosci M, Scarmeas N, Scarpini E, Scheltens P, Scherbaum N, Scherer M, Schmid M, Schneider A, Schott JM, Selbæk G, Seripa D, Serrano M, Sha J, Shadrin AA, Skrobot O, Slifer S, Snijders GJ, Soininen H, Solfrizzi V, Solomon A, Song Y, Sorbi S, Sotolongo-Grau O, Spalletta G, Spottke A, Squassina A, Stordal E, Tartan JP, Tárraga L, Tesí N, Thalamuthu A, Thomas T, Tosto G, Traykov L, Tremolizzo L, Tybjærg-Hansen A, Uitterlinden A, Ullgren A, Ulstein I, Valero S, Valladares O, Broeckhoven CV, Vance J, Vardarajan BN, van der Lugt A, Dongen JV, van Rooij J, van Swieten J, Vandenberghe R, Verhey F, Vidal JS, Vogelgsang J, Vyhnalek M, Wagner M, Wallon D, Wang LS, Wang R, Weinhold L, Wiltfang J, Windle G, Woods B, Yannakoulia M, Zare H, Zhao Y, Zhang X, Zhu C, Zulaica M, EADB, GR@ACE, DEGESCO, EADI, GERAD, Demgene, FinnGen, ADGC, CHARGE, Farrer LA, Psaty BM, Ghanbari M, Raj T, Sachdev P, Mather K, Jessen F, Ikram MA, de Mendonça A, Hort J, Tsolaki M, Pericak-Vance MA, Amouyel P, Williams J, Frikke-Schmidt R, Clarimon J, Deleuze JF, Rossi G, Seshadri S, Andreassen OA, Ingelsson M, Hiltunen M, Sleegers K, Schellenberg GD, van Duijn CM, Sims R, van der Flier WM, Ruiz A, Ramirez A, Lambert JC. New insights into the genetic etiology of Alzheimer's disease and related dementias. Nat Genet. 2022 Apr;54(4):412-436. Epub 2022 Apr 4 PubMed.
- Hemming ML, Selkoe DJ. Amyloid beta-protein is degraded by cellular angiotensin-converting enzyme (ACE) and elevated by an ACE inhibitor. J Biol Chem. 2005 Nov 11;280(45):37644-50. PubMed.
- Scotti L, Bassi L, Soranna D, Verde F, Silani V, Torsello A, Parati G, Zambon A. Association between renin-angiotensin-aldosterone system inhibitors and risk of dementia: A meta-analysis. Pharmacol Res. 2021 Apr;166:105515. Epub 2021 Feb 24 PubMed.
- Sink KM, Leng X, Williamson J, Kritchevsky SB, Yaffe K, Kuller L, Yasar S, Atkinson H, Robbins M, Psaty B, Goff DC. Angiotensin-converting enzyme inhibitors and cognitive decline in older adults with hypertension: results from the Cardiovascular Health Study. Arch Intern Med. 2009 Jul 13;169(13):1195-202. PubMed.
Further Reading
Papers
- Cao DY, Saito S, Veiras LC, Okwan-Duodu D, Bernstein EA, Giani JF, Bernstein KE, Khan Z. Role of angiotensin-converting enzyme in myeloid cell immune responses. Cell Mol Biol Lett. 2020;25:31. Epub 2020 May 25 PubMed.
Primary Papers
- Gomez AR, Byun HR, Wu S, Muhammad AK, Ikbariyeh J, Chen J, Muro A, Li L, Bernstein KE, Ainsworth R, Tourtellotte WG. Boosting angiotensin-converting enzyme (ACE) in microglia protects against Alzheimer's disease in 5xFAD mice. Nat Aging. 2025 Jul;5(7):1280-1294. Epub 2025 Jun 9 PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
Washington University in St. Louis
Washington University School of Medicine
While angiotensin-converting enzyme (ACE) is best known as a key component of the renin-angiotensin system and a crucial regulator of blood pressure, this study by Gomez et al. shows that its role extends far beyond this canonical function.
The authors focused on the role of ACE in the context of Alzheimer’s disease because it is one of the GWAS hits for late-onset AD. They generated a transgenic mouse line in which ACE is specifically overexpressed in microglia and examined its effect on amyloid pathology using the 5xFAD model. Their data show a protective effect of ACE overexpression, as evidenced by reduced plaque burden and neuronal loss, improved learning and memory, increased numbers of plaque-associated microglia, and enhanced Aβ phagocytosis by microglia. snRNA-Seq indicated an upregulation of Syk-mediated signaling pathways (PI3K/Akt, mTOR) in ACE-overexpressing disease-associated microglia (DAM) populations, and suppressing the Syk pathway in primary microglia by RNAi or pharmacological inhibition dampened their phagocytic ability toward oligomeric Aβ42. Overall, the study shows that ACE expression in microglia can boost their phagocytic function through Syk-mediated pathways, thereby constraining amyloid pathology. Although ACE is not expressed by microglia under homeostatic conditions or in AD, this work still offers valuable therapeutic insight, suggesting that introducing ACE-expressing microglial cells via cell-based therapy could be a potential strategy for treating AD.
Even though previous research has shown that ACE overexpression can reduce amyloid plaque burden in APP/PS1 mice (Bernstein et al., 2014), the underlying mechanism has remained unknown. This study is the first to comprehensively characterize the phenotypes and mechanisms associated with microglia-specific ACE overexpression. The authors also reported several findings that add novel insights to the microglia field, including the identification of the DAM-B population by snRNA-Seq, which has not been previously reported in amyloid models. Their spatial tissue microarray further showed that, unlike DAM-A (the canonical DAM population that cluster around plaques), the DAM-B population is located farther from plaques. It would be intriguing to understand what triggers the emergence of this unique DAM population and whether ACE contributes to its formation.
One of the missing links here is how ACE overexpression affects Syk signaling and alters microglia activation states. One possibility is through angiotensin II. Although angiotensin proteins in systemic circulation cannot cross the blood-brain barrier, astrocytes and neurons produce angiotensinogen, and the brain has its own local renin-angiotensin system (Labandeira-Garcia et al., 2017). Angiotensin II can act on AT1 or AT2 receptors, promoting or inhibiting inflammation and oxidative stress, respectively (Cosarderelioglu et al., 2020). An imbalance between these two pathways could contribute to neuroinflammation and alter microglial cellular states. Moreover, the phenotypes observed in this study could also be mediated by other ACE substrates, as ACE is a promiscuous peptidase that can even cleave Aβ peptides (Hemming and Selkoe, 2005) and convert Aβ1-42 to Aβ1-40 (Zou et al., 2007). It is possible that the excess ACE overexpressed by microglia facilitated Aβ degradation and partially contributed to the reduced pathology they observed.
References:
Bernstein KE, Gonzalez-Villalobos RA, Giani JF, Shah K, Bernstein E, Janjulia T, Koronyo Y, Shi PD, Koronyo-Hamaoui M, Fuchs S, Shen XZ. Angiotensin-converting enzyme overexpression in myelocytes enhances the immune response. Biol Chem. 2014 Oct;395(10):1173-8. PubMed.
Cosarderelioglu C, Nidadavolu LS, George CJ, Oh ES, Bennett DA, Walston JD, Abadir PM. Brain Renin-Angiotensin System at the Intersect of Physical and Cognitive Frailty. Front Neurosci. 2020;14:586314. Epub 2020 Sep 30 PubMed.
Hemming ML, Selkoe DJ. Amyloid beta-protein is degraded by cellular angiotensin-converting enzyme (ACE) and elevated by an ACE inhibitor. J Biol Chem. 2005 Nov 11;280(45):37644-50. PubMed.
Labandeira-Garcia JL, Rodríguez-Perez AI, Garrido-Gil P, Rodriguez-Pallares J, Lanciego JL, Guerra MJ. Brain Renin-Angiotensin System and Microglial Polarization: Implications for Aging and Neurodegeneration. Front Aging Neurosci. 2017;9:129. Epub 2017 May 3 PubMed.
Zou K, Yamaguchi H, Akatsu H, Sakamoto T, Ko M, Mizoguchi K, Gong JS, Yu W, Yamamoto T, Kosaka K, Yanagisawa K, Michikawa M. Angiotensin-converting enzyme converts amyloid beta-protein 1-42 (Abeta(1-42)) to Abeta(1-40), and its inhibition enhances brain Abeta deposition. J Neurosci. 2007 Aug 8;27(32):8628-35. PubMed.
Washington University School of Medicine
There has long been interest in understanding the role of ACE in Alzheimer’s disease. Studies have identified genetic variants in this gene that are associated with disease risk. Some studies suggest that ACE protein levels are lower in AD (Jochemsen et al., 2014) but other studies in larger datasets show higher levels in AD cases (Ali et al., 2025). In addition, it has been proposed that ACE inhibitors, which are normally used to treat blood pressure, reduce risk and AD progression, so it is not clear weather for AD, ACE needs to be activated or inhibited.
In this study, the authors overexpressed ACE, which is normally expressed in neurons, in microglia in APP/PS1 mice. They found that the ACE-expressed microglia increase phagocytosis and endolysosomal trafficking, reduce neuron and synapse degeneration, and even restore learning, which is quite promising. However, I would be cautious about the findings because it is not clear if they can be translated to humans, as the human-based data suggest that inhibiting rather than activating ACE may be beneficial for AD.
The manuscript indicated that expression of ACE in microglia increases microglial activity, which shows a protective effect. This is in line with increasing TREM2 activity, which also leads to more microglial activation. This raises the question about the timing of this potential intervention. Activating microglia in early stages of the disease may be beneficial, while in later stages it may be counterproductive. This duality could also explain the contradictory results seen in humans.
References:
Jochemsen HM, Teunissen CE, Ashby EL, van der Flier WM, Jones RE, Geerlings MI, Scheltens P, Kehoe PG, Muller M. The association of angiotensin-converting enzyme with biomarkers for Alzheimer's disease. Alzheimers Res Ther. 2014;6(3):27. Epub 2014 May 15 PubMed.
Ali M, Timsina J, Western D, Liu M, Beric A, Budde J, Do A, Heo G, Wang L, Gentsch J, Schindler SE, Morris JC, Holtzman DM, Ruiz A, Alvarez I, Aguilar M, Pastor P, Rutledge J, Oh H, Wilson EN, Guen YL, Khalid RR, Knight Alzheimer Disease Research Center (Knight ADRC), Alzheimer Disease Neuroimaging Initiative (ADNI), Fundació ACE Alzheimer Center Barcelona (FACE), Barcelona-1, Stanford Alzheimer Disease Research Center (Stanford ADRC), Robins C, Pulford DJ, Tarawneh R, Ibanez L, Wyss-Coray T, Sung YJ, Cruchaga C. Multi-cohort cerebrospinal fluid proteomics identifies robust molecular signatures across the Alzheimer disease continuum. Neuron. 2025 Mar 12; Epub 2025 Mar 12 PubMed.
Make a Comment
To make a comment you must login or register.