BACE and γ-Secretase Form Mega-Complex that Processes APP
Quick Links
The production of Aβ peptides from the amyloid precursor protein is a two-step affair: A first cut by the β-secretase (BACE) releases the extracellular portion of APP, leaving the C-terminal fragment (CTF) to find its way to γ-secretase for a final clipping to release Aβ. New data suggest the protease-to-protease handoff may be more intimate than previously thought. Work from the lab of Dennis Selkoe reveals that BACE, γ-secretase, and full-length APP can be isolated from mouse and human brain tissue in a high-molecular-weight complex that is capable of de novo production of Aβ peptides. The work, published January 9 in the Journal of Cell Biology, suggests the complex generates Aβ in vivo. A small molecule that destabilizes the union reduces Aβ production by cells, without directly inhibiting either BACE or γ-secretase catalytic activity.
- BACE and γ-secretase co-purify from mouse and human brain.
- Large complex processes full-length APP to Aβ peptides in vitro.
- Breaking it up may reduce Aβ while allowing processing of other substrates.
Stefan Lichtenthaler, German Center for Neurodegenerative Diseases, Munich, called the work an elegant follow-up of the lab’s previous studies on the interaction between α- and γ-secretases. “This is an important study with a very interesting concept, in that β-secretase forms a complex with γ-secretase that may allow easy proteolytic processing of substrate by both proteases,” he wrote to Alzforum. “Given the huge size of the complex, I am wondering whether β- and γ-secretase directly interact or whether they are co-purifying because they are in the same membrane microdomain, but without a direct interaction. I am sure the authors will investigate this further,” he wrote (see comment below).
“The presence of active multi-protease complexes is an exciting discovery,” said Gopal Thinakaran, University of Chicago. However, the location of the complexes, and their exact contribution to Aβ production in intact cells, remains to be clarified, he said.
To access γ-secretase, APP needs to shed its large extracelluar domain. That can be achieved by several membrane-bound proteases, most prominently BACE or α-secretase, also known as ADAM10. Previously, Selkoe’s lab discovered that α-secretase paired up with γ-secretase in large, multi-protease complexes that seemed to facilitate sequential proteolysis (Jan 2016 news). In the new study, first author Lei Liu asked if the same was true for BACE. Liu started with microsomes prepared from mouse brain, then solubilized the membrane proteins with mild detergent. In those preparations, antibodies to BACE1 consistently co-precipitated components of γ-secretase, including nicastrin and the N- and C-terminal fragments of presenilin1 (PS1). When Liu separated the solubilized membrane proteins using non-denaturing size-exclusion chromatography, he identified a complex of more than 5 million Daltons that harbored the γ-secretase proteins and full-length APP. Immunoprecipitation of this complex with anti-nicastrin antibodies pulled down BACE activity, suggesting the investigators had isolated a BACE1/γ-secretase complex analogous to their previously described α-secretase/γ-secretase pair. The complex contained only a fraction of the BACE in the cell: Most of the BACE1 protein, and protease activity, eluted in lower-molecular-weight fractions, without PS1.
Still, would the complex make Aβ? To measure this in vitro, scientists usually spike enzyme preps with recombinant APP. In this case, Liu took advantage of the endogenous APP already in the complex. He size-fractionated microsomes from HEK293 cells stably overexpressing human APP with the Swedish mutation, incubated the high-molecular-weight fractions at 37 degrees for 12 hours, solubilized them with detergent, and measured Aβ by ELISA. He found the complex had made Aβx-42 and Aβx-40 in a ratio of 0.1-0.2, which is within physiological norms. In contrast, the lower-molecular-weight fractions, with abundant BACE1 but no PS1, produced little or no Aβ. “This told us the BACE1/γ-secretase complex was functionally relevant, because the tiny amount of BACE1 in the high molecular weight fractions was, in fact, contributing to Aβ generation,” Liu told Alzforum. Complexes from cells expressing the familial AD PS1 mutants Y115H or L286V produced Aβ with a 42/40 ratio of 0.5 or higher, supporting the idea that these complexes are a physiological source of the peptides. Based on the cell fractionation experiments, Selkoe thinks most of the Aβ in cells could be derived from the high-molecular-weight BACE1/γ-secretase assemblies.
Surprisingly, the γ-secretase active site inhibitor L685,458 did not diminish Aβ production by the high-molecular-weight complex, though L685,458 suppresses Aβ production in human neuronal cells (Tagami et al., 2017). Liu and Selkoe speculated it had no effect on the complexes because APP was already bound and prevented the inhibitor from accessing the PS1 active site. A PS1 allosteric modulator that binds outside the catalytic site did have the expected effect: JNJ-40418677 attenuated production of Aβx-42 by the complex, and boosted Aβx-37.
To prove the complex sequentially cleaved full-length APP, and not simply APP CTFs bound in the complex, the investigators depleted CTFs in HEK293-swAPP cells by treatment with the BACE inhibitor AZD3293. This increased the amount of full-length APP in the cell and when Liu isolated the high-molecular-weight BACE/γ-secretase, now sans AZD3293, it made 1.5- to fourfold more Aβ than complexes from untreated cells.
Can the same complex be isolated from human brain? Apparently, yes. Liu size-fractionated preparations of fresh human brain tissue, and showed the HMW fractions produced the bulk of Aβ from endogenous APP. Aβ production by the complex was only marginally inhibited by three different γ-secretase inhibitors, but, as they saw with HEK293 cells, the modulator JNJ-40418677 shifted cleavage toward the smaller Aβx-37.
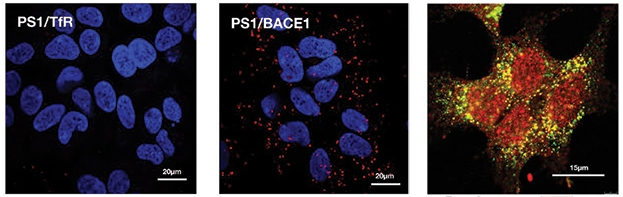
Close companions. Proximity ligation (red puncta) links PS1 to BACE1 (middle panel), but not to a transferrin receptor control (left panel). Super-resolution STED microscopy shows PS1 (red) and BACE1 (green) in individual puncta (right). [Courtesy of Liu et al., JCB 2019.]
The scientists next asked if they could target the complex to reduce Aβ production as a potential therapeutic strategy. Previously, Chinese researchers had discovered that the natural product 3-α-akebonoic acid (3AA) disrupted BACE/γ-secretase interactions in cells and crippled Aβ secretion (Cui et al., 2015). Liu showed that a related compound, roburic acid, inhibited Aβ production from HEK293-swAPP cells. This was tied to reduced γ-secretase and BACE activity in the large complex. Roburic acid did not directly inhibit BACE1 or γ-secretase enzymatic activity, although it did show some γ-secretase modulatory activity, decreasing the Aβx-42/x-40 ratio and increasing the Aβx-38/x-42 ratio. “Roburic acid or other compounds that separate the BACE and γ-secretases, without affecting their active sites, could offer an alternative approach to BACE inhibitors,” Selkoe said. With BACE-inhibitor clinical trials turning up discouraging cognitive and other side effects, possibly due to decreased processing of other important substrates, new approaches are clearly needed, he added (Nov 2018 conference news).
Given that BACE and γ-secretase travel in different circles in the cell, with distinct targeting and trafficking pathways, Thinakaran wondered where they would meet up to form a large complex. “The next challenges are to determine where within the neurons the multi-protease α/γ and β/γ complexes are assembled, and characterize the trafficking motifs in the individual components of the complexes that cooperate or act as predominant signals to direct the complexes to their subcellular sites within neurons,” he said. By several methods, including proximity ligation (see image above), Selkoe’s group detected close association of BACE1 and γ-secretase near nuclei of HEK cells. Selkoe said their techniques establish that the proteins are together, but they do not yet know precisely where they sit in the cell. "We believe there must be physical loci where this complex resides, but we’re not sure where that is yet,” he said.
“It is absolutely conceivable that the secretase activities act to process or degrade substrates such as APP in a concerted manner,” said Stephan Schilling, Fraunhofer Institute for Cell Therapy and Immunology, Leipzig, Germany. “Indeed, such ‘substrate tunneling’ is a conserved biochemical principle, which is, for example, known for enzymes acting in amino-acid synthesis in the cytosol. The principle is that products of one processing step are not released into bulk but are rather guided to another enzyme within a complex, which makes these processes much more efficient. That this principle might also apply to membrane processing by proteases is an intriguing finding, and the authors collected compelling evidence for formation of such a multi-enzyme complex,” he wrote to Alzforum.
Liu spied another potential APP sheddase in the high-molecular-weight fraction. The metalloprotease meprin β has been proposed as an alternative BACE that cleaves APP at the same site and thus might also feed presenilin (Becker-Pauly and Pietrzik, 2016). “The localization of meprin β to HMW fractions containing APP and γ-secretase provides another important piece of evidence that this enzyme contributes to Aβ generation in the brain,” wrote Claus Pietrzik, Johannes Gutenberg University Mainz, Germany, to Alzforum. However, Liu said the presence of the enzyme does not mean it’s necessarily a partner of γ-secretase. They have not yet done the co-immunoprecipitation experiments to determine that.—Pat McCaffrey
References
News Citations
Mutations Citations
Paper Citations
- Tagami S, Yanagida K, Kodama TS, Takami M, Mizuta N, Oyama H, Nishitomi K, Chiu YW, Okamoto T, Ikeuchi T, Sakaguchi G, Kudo T, Matsuura Y, Fukumori A, Takeda M, Ihara Y, Okochi M. Semagacestat Is a Pseudo-Inhibitor of γ-Secretase. Cell Rep. 2017 Oct 3;21(1):259-273. PubMed.
- Cui J, Wang X, Li X, Wang X, Zhang C, Li W, Zhang Y, Gu H, Xie X, Nan F, Zhao J, Pei G. Targeting the γ-/β-secretase interaction reduces β-amyloid generation and ameliorates Alzheimer's disease-related pathogenesis. Cell Discov. 2015;1:15021. Epub 2015 Aug 18 PubMed.
- Becker-Pauly C, Pietrzik CU. The Metalloprotease Meprin β Is an Alternative β-Secretase of APP. Front Mol Neurosci. 2016;9:159. Epub 2017 Jan 5 PubMed.
Further Reading
No Available Further Reading
Primary Papers
- Liu L, Ding L, Rovere M, Wolfe MS, Selkoe DJ. A cellular complex of BACE1 and γ-secretase sequentially generates Aβ from its full-length precursor. J Cell Biol. 2019 Feb 4;218(2):644-663. Epub 2019 Jan 9 PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
K.U.Leuven and V.I.B.
The co-localization of β-secretase, γ-secretase, and APP in ~5MDa high-molecular-weight complexes is clear and indeed raises the possibility of a coupled regulated intermembrane proteolysis (RIP) process. Interestingly, confocal microscopy (proximity ligation analysis-data) evidences a short distance between the secretases (< 40 nM), suggesting a direct interaction. Whether this comes with functional consequences is less clear. The final proof will likely require the reconstitution of the β-secretase/γ-secretase/APP complex and functional/structural analyses.
It would be very exciting to look at the tripartite complex at atomic resolution!
German Center for Neurodegenerative Diseases (DZNE)
This is an elegant follow-up of the authors’ previous paper on the interaction between α- and γ-secretase. This is an important study with a very interesting concept, in that β-secretase forms a complex with γ-secretase, which may allow easy proteolytic processing of a substrate by both proteases. Given the huge size of the complex, I am wondering whether β- and γ-secretase directly interact or whether they are co-purifying because they are in the same membrane microdomain, but without a direct interaction. I am sure the authors will investigate this further and may then come up with more proteins being part of such a complex. Using mass spec on the purified complexes may be one way to go.
University of Connecticut Health
It is a very interesting study, and does provide a logical explanation of how Aβ can be efficiently processed by both β- and γ-secretase in a large complex. One intriguing question is how many such high molecular weight (HMW) complexes will exist, because most BACE1 substrates, such as Neuregulin-1, can also be processed by γ-secretase. How stable will such a HMW complex be in cells or brains?
Johannes Gutenberg University Mainz
This carefully conducted study demonstrates that Aβ generation in the brain occurs in an endogenous high-molecular-weight complex (HMW) containing the substrate full-length APP and proteolytically active β- and γ-secretases. In this endogenous complex, which was identified by Liu and colleges using non-denaturing FPLC combined with novel Aβ ELISAs in wild-type mouse and human brain, APP undergoes sequential β- and γ-secretase cleavages. In additional in vitro studies with HEK cells overexpressing Swedish-mutant APP, Liu and colleagues provide further evidence that BACE1 and γ-secretase are the most important enzymes to generate Aβ peptides from this FAD mutation. In general, these HMW fractions could be very useful tools to test and optimize secretase inhibitors and modulators and to further develop safe and effective AD drugs.
Although BACE1 most likely acts as the major β-secretase, several N-terminally truncated Aβ variants including peptides starting at position p2 have been described in the CSF and brains of sporadic AD patients and cannot be attributed to BACE 1 activity (Wiltfang et al., 2001; Bibl et al., 2012).
I am intrigued that Liu and colleagues were able to demonstrate the presence of the metalloprotease meprin β in the same mouse brain HMW fractions containing holo-APP, BACE1, and γ-secretase. We have shown that meprin β is able to generate N-terminally truncated Aβ-peptides starting at position p2 (Bien et al., 2012), and that this only occurs when wild-type and not Swedish-mutant APP is expressed (Schoenherr et al., 2016). Additionally, Schlenzig and colleagues have recently demonstrated an increase in meprin β expression in AD brains (Schlenzig et al., 2018). Nevertheless, so far, proof that endogenously expressed meprin β plays a role in the generation of N-terminal truncated Aβ2-x peptides in vivo or in AD is still missing. Therefore, we suggest that the localization of meprin β to HMW fractions containing APP and γ-secretase provides another important piece of evidence that this enzyme contributes to Aβ generation in the brain.
References:
Bien J, Jefferson T, Causevic M, Jumpertz T, Munter L, Multhaup G, Weggen S, Becker-Pauly C, Pietrzik CU. The Metalloprotease Meprin β Generates Amino Terminal-truncated Amyloid β Peptide Species. J Biol Chem. 2012 Sep 28;287(40):33304-13. PubMed.
Schönherr C, Bien J, Isbert S, Wichert R, Prox J, Altmeppen H, Kumar S, Walter J, Lichtenthaler SF, Weggen S, Glatzel M, Becker-Pauly C, Pietrzik CU. Generation of aggregation prone N-terminally truncated amyloid β peptides by meprin β depends on the sequence specificity at the cleavage site. Mol Neurodegener. 2016 Feb 19;11:19. PubMed.
Schlenzig D, Cynis H, Hartlage-Rübsamen M, Zeitschel U, Menge K, Fothe A, Ramsbeck D, Spahn C, Wermann M, Roßner S, Buchholz M, Schilling S, Demuth HU. Dipeptidyl-Peptidase Activity of Meprin β Links N-truncation of Aβ with Glutaminyl Cyclase-Catalyzed pGlu-Aβ Formation. J Alzheimers Dis. 2018;66(1):359-375. PubMed.
Wiltfang J, Esselmann H, Cupers P, Neumann M, Kretzschmar H, Beyermann M, Schleuder D, Jahn H, Rüther E, Kornhuber J, Annaert W, De Strooper B, Saftig P. Elevation of beta-amyloid peptide 2-42 in sporadic and familial Alzheimer's disease and its generation in PS1 knockout cells. J Biol Chem. 2001 Nov 16;276(46):42645-57. PubMed.
Bibl M, Gallus M, Welge V, Esselmann H, Wolf S, Rüther E, Wiltfang J. Cerebrospinal fluid amyloid-β 2-42 is decreased in Alzheimer's, but not in frontotemporal dementia. J Neural Transm. 2012 Jul;119(7):805-13. PubMed.
Fraunhofer Institute for Cell Therapy and Immunology
This is really interesting work. From a general viewpoint, it appears absolutely conceivable to me that the secretase activities act to process or degrade substrates such as APP in a concerted manner. Indeed, such “substrate tunneling” is a conserved biochemical principle, which is, e.g., known for enzymes acting in amino acid synthesis in the cytosol. The principle is that products of one processing step are not released into bulk but rather are guided to another enzyme within a complex, which makes these processes much more efficient.
That this principle might also apply to membrane processing by proteases is an intriguing finding, and the authors collected compelling evidence for formation of such a multi-enzyme-complex. Unfortunately, the authors do not comment on meprin β in the complex. Our data and that from other labs support APP cleavage by this protease. It would have been interesting to study the N-terminus of the Aβ peptides, but this was certainly beyond the scope of the study.
Tongji University/Chinese Academy of Sciences
Congratulations to Dr. Selkoe's lab for their wonderful work! I am really impressed by their delicate and systematic approaches to convincingly demonstrate the β- and γ-secretase complex and its unique catalytic activity.
In our earlier discovery (Cui et al., 2015), we found that β- and γ-secretases indeed functionally interacted with each other to carry out the well-known sequential cleavage of APP. Though in some of our experiments, such as co-localization and fractioning, the complex of β- and γ-secretases (also with other partners) was observed, our efforts then focused on proving the concept that their interaction could be a potential drug target for disrupting Aβ generation and for AD therapy. Through high-throughput screening and chemical modification, 3-α-akebonoic acid (3AA) and its derivatives were identified as effectively reducing Aβ production and further as alleviating cognitive dysfunction and Aβ-related pathology in an AD mouse model.
In this wonderful work by Liu et al., the complex containing β- and γ-secretases was not only identified in cultured cells but also in mouse and human brain, and the complex’s catalytic ability to generate a full array of Aβ peptides was compellingly shown in vitro. This might serve as a convenient assay for further large-scale screening and more mechanistic study. Most interesting, they found that Roburic acid, an analogue of 3-α-akebonoic acid, dose-dependently reduced Aβ production by interfering with β-/γ-secretase complexes and modulating γ-secretase. Their results really encourage and also facilitate us to continue our exploration for better and more druggable disruptor candidates.
It may be worth pointing out here that in our recent research we found that α-secretase (ADAM10) physically interacted with BACE1 in neurons with some proteolytic regulatory function (Wang et al., 2018). Further study by fluorescence resonance energy transfer showed that α-/β-/γ-secretase likely exists in a ternary complex (even with more partners) (Wang and Pei, 2018), suggesting that α- and β-secretases may compete adjacent each other and dynamically, with physical and functional involvement of γ-secretase, APP, and other stakeholders. We hope that by those delicate methodologies in Dr. Selkoe's work and by the up-to-date Cryo-electron microscopy, the structure of the complexes will be solved soon, and the underlying mechanisms of various modifiers such as genetic, epigenetic, environmental, as well as aging would be better understood.
Aging seems inevitable for living creatures, including ourselves, but AD could be prevented or at least delayed. The disappointing outcome of AD treatment for the last decades probably teaches us to study more and to try alternative and more creative approaches.
Make a Comment
To make a comment you must login or register.