At the Heart of Every ALS RNA Aggregate, a Single Transcript
Quick Links
Expanded hexanucleotide repeats in the C9ORF72 gene cause more cases of familial amyotrophic lateral sclerosis and frontotemporal dementia than any other genetic risk factor. Mutant C9ORF72 RNA accumulates in aggregates that may play a role in toxicity, but little is known about their composition. Now, in the January 18 Cell Chemical Biology, researchers led by David Corey at the University of Texas Southwestern Medical Center, Dallas, report that cells from people with the expansion express surprisingly few copies of the mutant gene, averaging only one to three RNAs per cell. This matched the average number of visible RNA foci per cell. This implies that a single expanded RNA molecule lies at the core of each deposit. Since the amount of mutant transcript determines how much protein can be made or sequestered by the RNA, the finding could provide clues to disease pathogenesis, Corey told Alzforum. “I hope more scientists will count RNA molecules. You can’t understand disease mechanisms unless you know what the numbers are,” Corey said. In future work, he plans to expand this type of analysis to other repeat diseases.
Johnathan Cooper-Knock at the University of Sheffield, U.K., expressed enthusiasm for the finding, though he noted it needs to be repeated in postmortem tissue to see if the phenomenon holds true in the brain. “This is really interesting, and leads to important ideas. If the authors are right, and we are talking about very small numbers of molecules per cell, then the repeat length should be closely related to disease severity,” he suggested.
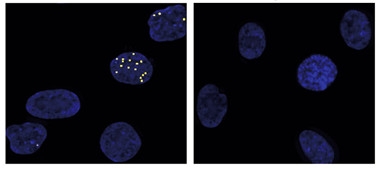
One Per Dot.
RNA foci (white) accumulate in nuclei (blue) of fibroblasts carrying expanded C9ORF72 (left), but not in those from controls. [Courtesy of Cell Chemical Biology, Liu et al.]
Patients with the C9ORF72 mutation carry massive expansions of up to 1,600 copies of a GGGGCC hexamer repeat in the first intron of the gene, compared to 23 or fewer repeats in unaffected people (see Sep 2011 news). The expanded gene is transcribed in both directions, leading to deposits of sense and antisense RNA, but data conflict on whether these RNA foci are toxic. Some animal studies report that mice and flies function fine despite RNA accumulation, whereas induced cells and brain tissue from patients show evidence of toxicity, particularly for antisense foci, which accumulate TDP-43 (see Sep 2015 news; Dec 2015 news; Oct 2013 news; May 2015 news).
None of the previous studies tried to count the number of RNA molecules, however. The number matters, because that provides clues to gene regulation and disease mechanisms, Corey noted (see Bosson et al., 2014; Denzler et al., 2014). To quantify the RNA, first author Jing Liu took cultures of fibroblasts donated by patients heterozygous for the expansion and by healthy controls, isolated C9ORF72 RNA from them, and generated cDNA from it. Since each mRNA only generates one cDNA with their method, the authors were able to estimate transcript number by counting C9ORF72 cDNA molecules. They did so using two independent methods, quantitative PCR and droplet digital PCR. They divided the total number of cDNA molecules by the number of cells in the sample to obtain the average copy number per cell. Different primers allowed them to distinguish between mature messenger RNA, from which the first intron had been excised, and immature sense and antisense transcripts containing the expanded intron 1 repeats.
Both PCR methods produced similar results. Fibroblasts from patients contained 15 to 20 copies of mature C9ORF72 RNA per cell, and one to three copies each of immature sense and antisense expanded RNAs. Less transcription occurred in control cells, which averaged five to 10 copies of the mature C9ORF72 RNA, along with one sense and one antisense immature—but not expanded—transcript per cell. Nearly all of the expanded patient RNAs remained in the nucleus, while about one-third of the mature RNA message made it to the cytoplasm.
This is the key experiment: The authors compared the RNA copy number to the amount of foci per cell, as determined by fluorescence in situ hybridization (FISH). They found a close correspondence, with patient cells averaging one focus per expanded RNA. However, the number of deposits per cell varied. About 60 percent of patient-derived fibroblasts had none, while 1 to 2 percent of cells accumulated more than 10 each (see image above). This pattern agrees with previous findings in fibroblasts and induced cells, by the Corey lab and others (see Oct 2013 news; Hu et al., 2015). Perhaps the handful of cells that contain numerous foci play a disproportionate role in disease, Corey speculated.
Would neurons and glia make similar numbers of transcripts? The authors generated induced pluripotent stem cells from patients who carried one regular and one expanded C9ORF72 gene, and then differentiated them into neurons and astrocytes. Like fibroblasts, astrocytes carrying the mutation harbored about 12 copies of the mature RNA and one copy each of the sense and antisense expanded C9ORF72 RNAs. In motor neurons with the mutation, however, antisense transcripts dominated. These cells made about one copy of the sense RNA, but 10 of the antisense version. Control neurons, by contrast, made only one of each. They did not examine the transcript:foci ratio in these induced cells.
Intriguingly, motor neurons in postmortem tissue from C9ORF72 mutation carriers also harbor an excess of mutant antisense RNA compared to sense (see Cooper-Knock et al., 2015). Moreover, in a recent C9ORF72 mouse model generated by Laura Ranum at the University of Florida, Gainesville, degenerating brain areas contained more antisense than sense RNA, again suggesting this form might be particularly toxic (see May 2016 news).
Whether the finding of one RNA per focus holds in vivo remains unclear. Individual motor neurons from postmortem brains contain multiple foci of different sizes, suggesting that some contain more RNA than others, Cooper-Knock noted. RNA foci attract proteins as well, though the amount of protein they can bind remains proportional to the RNA length, Corey said.
Cooper-Knock found the most exciting implication of the work to be that it reinforces the assumed relationship between repeat length and toxicity. With so few copies of mutant RNA per cell, the size of the expansion determines how much expanded repeat RNA is present in each cell to wreak havoc, not the number of transcripts, he pointed out. Previous research found only a rough correlation between repeat length and disease (see van Blitterswijk et al., 2015). However, these older studies were based on DNA length, not RNA. They used Southern blots, which are not very accurate, Cooper-Knock said. It would be interesting to examine patient samples with newer sequencing techniques to see if repeat length does forecast severity, he said.
That there are so few transcripts per cell further suggests that antisense therapeutic strategies could be particularly effective in shutting down transcription, researchers agreed. These approaches have recently garnered an FDA approval for spinal muscular atrophy (see Nov 2016 news), and are in Phase 1 for Huntington’s disease, and Phase 1 for ALS (see Dec 2015 conference news).—Madolyn Bowman Rogers
References
News Citations
- Corrupt Code: DNA Repeats Are Common Cause for ALS and FTD
- C9ORF72 RNA Foci Acquitted of Toxic Charge—in Fruit Flies
- C9ORF72 Mice A-OK Despite Toxic RNAs, Peptides
- RNA Deposits Confer Toxicity in C9ORF72 ALS
- Antisense RNA from C9ORF72 Repeats Is Likely Culprit in Patient Neurons
- Second Study Confirms Antisense Oligonucleotides Bust RNA Aggregates
- New C9ORF72 Mice Develop Symptoms Resembling ALS/FTD
- Positive Trials of Spinal Muscular Atrophy Bode Well for Antisense Approach
- Gene and Stem Cell Therapies Make Strong Showing at ALS/MND Meeting
Paper Citations
- Bosson AD, Zamudio JR, Sharp PA. Endogenous miRNA and target concentrations determine susceptibility to potential ceRNA competition. Mol Cell. 2014 Nov 6;56(3):347-59. Epub 2014 Oct 23 PubMed.
- Denzler R, Agarwal V, Stefano J, Bartel DP, Stoffel M. Assessing the ceRNA hypothesis with quantitative measurements of miRNA and target abundance. Mol Cell. 2014 Jun 5;54(5):766-76. Epub 2014 May 1 PubMed.
- Hu J, Liu J, Li L, Gagnon KT, Corey DR. Engineering Duplex RNAs for Challenging Targets: Recognition of GGGGCC/CCCCGG Repeats at the ALS/FTD C9orf72 Locus. Chem Biol. 2015 Nov 19;22(11):1505-11. Epub 2015 Nov 12 PubMed.
- Cooper-Knock J, Higginbottom A, Stopford MJ, Highley JR, Ince PG, Wharton SB, Pickering-Brown S, Kirby J, Hautbergue GM, Shaw PJ. Antisense RNA foci in the motor neurons of C9ORF72-ALS patients are associated with TDP-43 proteinopathy. Acta Neuropathol. 2015 Jul;130(1):63-75. Epub 2015 May 6 PubMed.
- van Blitterswijk M, Gendron TF, Baker MC, DeJesus-Hernandez M, Finch NA, Brown PH, Daughrity LM, Murray ME, Heckman MG, Jiang J, Lagier-Tourenne C, Edbauer D, Cleveland DW, Josephs KA, Parisi JE, Knopman DS, Petersen RC, Petrucelli L, Boeve BF, Graff-Radford NR, Boylan KB, Dickson DW, Rademakers R. Novel clinical associations with specific C9ORF72 transcripts in patients with repeat expansions in C9ORF72. Acta Neuropathol. 2015 Dec;130(6):863-76. Epub 2015 Oct 5 PubMed.
Further Reading
Primary Papers
- Liu J, Hu J, Ludlow AT, Pham JT, Shay JW, Rothstein JD, Corey DR. c9orf72 Disease-Related Foci Are Each Composed of One Mutant Expanded Repeat RNA. Cell Chem Biol. 2017 Feb 16;24(2):141-148. Epub 2017 Jan 26 PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
No Available Comments
Make a Comment
To make a comment you must login or register.