Do Microglia and Astrocytes Fuel the Alzheimer’s Cascade?
Quick Links
Move over, amyloid, reactive astrocytes and activated microglia may be the main drivers of the Alzheimer’s disease pathological cascade. In the November 6 Nature Neuroscience, scientists led by Pedro Rosa-Neto of McGill University, Montreal, Canada, and Eduardo Zimmer of the Universidade Federal do Rio Grande do Sul, Porto Alegre, Brazil, report that in the human brain, amyloid stirs up astrocyte reactivity—but only in the presence of activated microglia. In turn, this riled-up duo unleashes tau pathology and cognitive decline, pointing to microglia–astrocyte crosstalk as a key link in the cellular phase of AD.
- In Alzheimer’s disease, astrocyte reactivity rises with amyloid.
- But only when microglia are activated.
- When both glial cells are riled, tau tangles and cognitive decline follow.
“This fascinating work reveals that the glial response seen in Alzheimer’s disease is not merely a passive reaction to Aβ buildup, but an active, multi-step process in which crosstalk between microglia and astrocytes directly impacts neuronal health and cognitive function,” Amaia Arranz Mendiguren of the Achucarro Basque Center for Neuroscience in Leioa, Spain, told Alzforum (comment below).
Previous work led by Tharick Pascoal at the University of Pittsburgh, in collaboration with Zimmer, Rosa-Neto, and several co-authors of the present study, found that among individuals with brain amyloid, only those with elevated astrocyte reactivity—measured by plasma glial fibrillary acidic protein (GFAP)—also had tau pathology. Indeed, baseline amyloid predicted tangle spread only in participants with high GFAP, nominating astrocyte reactivity as an important intermediary between amyloid and tau (Bellaver et al., 2023; Jun 2023 news). How do microglia fit in?
To find out, first authors João Pedro Ferrari-Souza in Zimmer’s lab, Guilherme Povala at U Pitt, and Nesrine Rahmouni at McGill analyzed data from participants aged 50 and older in the Translational Biomarkers in Aging and Dementia (TRIAD) cohort, a biomarker study that began enrolling volunteers in 2017 at the McGill University Research Centre for Studies in Aging. The researchers included 62 cognitively unimpaired participants, 26 with mild cognitive impairment, and 13 with Alzheimer’s disease. They used [18F]AZD4694 PET imaging to measure Aβ and [11C]PBR28 PET to measure translocator protein expressed on the surface of activated microglia. Because a common TSPO polymorphism—an alanine-to-threonine substitution at position 147—affects how tightly [11C]PBR28 binds, the researchers genotyped participants and included only those with the high-affinity alanine/alanine form, found in about 50 percent of the cohort. They also measured plasma GFAP and p-tau217 levels.
The scientists divided participants into those testing positive or negative on TSPO PET. In the former, plasma GFAP levels rose in lockstep with both amyloid PET signal and plasma p-tau217. These relationships crumbled in TSPO–negative participants (image below).

Microglia Matter. In TSPO-positive participants (black), plasma GFAP rose in tandem with amyloid (left) and p-tau217 (right). In TSPO–negative participants (tan), this correlation was negligible. [Courtesy of Ferrari-Souza et al., Nature Neuroscience, 2025.]
TSPO imaging has come under fire in recent years, since its signal can arise from astrocytes and endothelial cells, and may even reflect microglial density rather than activation (Lavisse et al., 2012; May 2022 news; Nutma et al., 2023). For an alternate marker, the researchers used soluble TREM2, which sloughs off proinflammatory microglia. They measured sTREM2 in the cerebrospinal fluid of participants from the TRIAD and the independent Wisconsin Registry for Alzheimer’s Prevention (WRAP) cohorts. WRAP, based at the University of Wisconsin School of Medicine and Public Health, Madison, has followed cognitively normal people with a parental history of Alzheimer’s since 2001. Ferrari-Souza limited analysis to unimpaired and mildly impaired participants in both groups. Among the 254 participants, of whom 37 were mildly impaired, amyloid burden and astrocyte reactivity appeared intertwined. In those with the highest CSF sTREM2, however, the correlation was nearly three times stronger, suggesting that amyloid exerts a much stronger effect on astrocytes when microglia are activated.
What do these correlations say about AD progression? To find out, the scientists used structural equation modeling, a statistical approach that tests relationships among multiple variables. In participants with no microglial activation, higher amyloid load associated with elevated plasma p-tau217 and greater tau pathology on PET, but not astrocyte activation or cognitive impairment. In those with high microglial activation, the pieces fit neatly together: amyloid load, astrocyte reactivity, tau accumulation, and cognition were all statistically linked (image below).
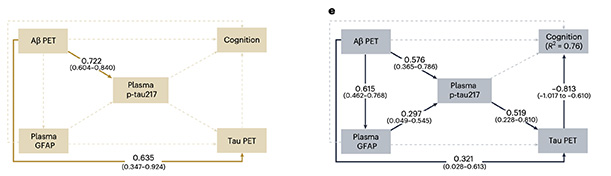
Complete the Circuit. In participants with no microglial activation (left), statistical modelling revealed strong links between amyloid burden, plasma p-tau217, and tangle pathology, but little association with astrocyte activity or cognitive decline. When microglia are activated (right), astrocyte reactivity enters the mix, tangle pathology worsens, and cognition falters. [Courtesy of Ferrari-Souza et al., Nature Neuroscience, 2025.]
“The interconnection between microglia and astrocytes has been widely studied in vitro and in animal models, but being able to observe this relationship in humans, in vivo, is exciting,” noted Andrea Lessa Benedet of the University of Gothenburg, Sweden. She is a co-author. Other scientists cautioned that the data are correlative and do not confirm a direct, Aβ-specific interaction between microglia and astrocytes.
The underlying mechanisms seem complex, since both types of glia can exhibit neuroprotective and neurotoxic effects depending on disease stage, noted Agneta Nordberg of the Karolinska Institute in Sweden. “The exact mechanism by which microglia, as discussed in the present paper, may modulate Aβ-induced astrocyte reactivity is still unknown,” she said.
Rosa-Neto plans to address the mechanism in future work. Besides expanding their PET imaging readouts, his team will analyze CSF to measure inflammatory mediators and gene expression in immune cells from individuals with varying degrees of Alzheimer’s pathology.—George R. Heaton
George Heaton is a freelance writer in Durham, North Carolina.
References
News Citations
- In Amyloid Cascade, Do Reactive Astrocytes Bridge Plaques and Tangles?
- Does TSPO PET Measure Microglia Activation or Density?
Paper Citations
- Bellaver B, Povala G, Ferreira PC, Ferrari-Souza JP, Leffa DT, Lussier FZ, Benedet AL, Ashton NJ, Triana-Baltzer G, Kolb HC, Tissot C, Therriault J, Servaes S, Stevenson J, Rahmouni N, Lopez OL, Tudorascu DL, Villemagne VL, Ikonomovic MD, Gauthier S, Zimmer ER, Zetterberg H, Blennow K, Aizenstein HJ, Klunk WE, Snitz BE, Maki P, Thurston RC, Cohen AD, Ganguli M, Karikari TK, Rosa-Neto P, Pascoal TA. Astrocyte reactivity influences amyloid-β effects on tau pathology in preclinical Alzheimer's disease. Nat Med. 2023 Jul;29(7):1775-1781. Epub 2023 May 29 PubMed.
- Lavisse S, Guillermier M, Hérard AS, Petit F, Delahaye M, Van Camp N, Ben Haim L, Lebon V, Remy P, Dollé F, Delzescaux T, Bonvento G, Hantraye P, Escartin C. Reactive astrocytes overexpress TSPO and are detected by TSPO positron emission tomography imaging. J Neurosci. 2012 Aug 8;32(32):10809-18. PubMed.
- Nutma E, Fancy N, Weinert M, Tsartsalis S, Marzin MC, Muirhead RC, Falk I, Breur M, de Bruin J, Hollaus D, Pieterman R, Anink J, Story D, Chandran S, Tang J, Trolese MC, Saito T, Saido TC, Wiltshire KH, Beltran-Lobo P, Phillips A, Antel J, Healy L, Dorion MF, Galloway DA, Benoit RY, Amossé Q, Ceyzériat K, Badina AM, Kövari E, Bendotti C, Aronica E, Radulescu CI, Wong JH, Barron AM, Smith AM, Barnes SJ, Hampton DW, van der Valk P, Jacobson S, Howell OW, Baker D, Kipp M, Kaddatz H, Tournier BB, Millet P, Matthews PM, Moore CS, Amor S, Owen DR. Translocator protein is a marker of activated microglia in rodent models but not human neurodegenerative diseases. Nat Commun. 2023 Aug 28;14(1):5247. PubMed.
Further Reading
Papers
- Liddelow SA, Guttenplan KA, Clarke LE, Bennett FC, Bohlen CJ, Schirmer L, Bennett ML, Münch AE, Chung WS, Peterson TC, Wilton DK, Frouin A, Napier BA, Panicker N, Kumar M, Buckwalter MS, Rowitch DH, Dawson VL, Dawson TM, Stevens B, Barres BA. Neurotoxic reactive astrocytes are induced by activated microglia. Nature. 2017 Jan 26;541(7638):481-487. Epub 2017 Jan 18 PubMed.
Primary Papers
- Ferrari-Souza JP, Povala G, Rahmouni N, Bellaver B, Ferreira PC, De Bastiani MA, Leffa DT, Lussier FZ, Aguzzoli CS, Brum WS, Carello-Collar G, Borelli WV, Therriault J, Macedo AC, Servaes S, Stevenson J, Pola I, Gauthier S, Souza DO, Schilling LP, Lourenco MV, Triana-Baltzer G, Kolb HC, Benedet AL, Ashton NJ, Tudorascu DL, Zetterberg H, Blennow K, Johnson SC, Pascoal TA, Rosa-Neto P, Zimmer ER. Microglia modulate Aβ-dependent astrocyte reactivity in Alzheimer's disease. Nat Neurosci. 2025 Nov 6; Epub 2025 Nov 6 PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
Achucarro Basque Center for Neuroscience
This fascinating work by Eduardo Zimmer and colleagues reveals that the glial response observed in Alzheimer's disease is not just a passive reaction to Aβ accumulation, but an active, multi-step process in which the crosstalk between microglia and astrocytes has direct consequences for neuronal health and cognitive function. Remarkably, the use of clinical biomarkers such as TSPO (microglial activation PET) and GFAP (astrocyte reactivity plasma biomarker) provide crucial clinical evidence indicating that these interactions between microglia and astrocytes are relevant to human disease progression. Moreover, this work also underscores that targeting this interplay between microglia and astrocytes, rather than just Aβ or tau directly, might be a critical therapeutic strategy in Alzheimer's disease.
University of Gothenburg
What João Pedro has reported is in line with what I believe happens in the disease: an inflammatory trigger provides a major push driving the disease forward, and astrocytic reactivity, or dysfunction, represents an important component of the AD pathophysiological process, together with amyloid accumulation, tau pathology, and synaptic degeneration.
The interconnection between microglia and astrocytes has been widely studied in vitro and in animal models, but being able to observe this relationship in humans, in vivo, is exciting, and gives us much to think about. What makes one’s immune response more prone to triggering these negative downstream effects? An accumulation of stressors? Genetic factors? Disease burden? All of the above?
We also don’t yet fully understand what plasma GFAP truly represents in terms of pathology. GFAP is not equally expressed by astrocytes across brain regions, and not all activated astrocytes express GFAP. This remains an important knowledge gap, and filling it would help us better interpret the current findings.
Karolinska Institutet
It is apparent that both microglia and astrocytes are key players in the neuroinflammatory response observed in Alzheimer´s disease AD. The underlying mechanisms seem still to be complex, since both types of glia cells can exhibit neuroprotective and neurotoxic mechanisms depending on disease stage. This paper focuses on how microglia can modulate the well-known Aß-dependent astrocyte reactivity in AD.
Microglia PET with [ 11C]PBR28 required PET scans of 606 individuals in order to obtain 101 high-affinity bind-ers, and the microglia activation is only 9.6 % higher in cognitively impaired (n=39) than cognitively unimpaired (n=62) participants in the study. It would have been val-uable if, in addition to plasma GFAP levels, reactive as-trogliosis had also been measured by PET, since these two biomarkers seem to follow divergent trajectories in AD (Chiotis et al. 2023).
Increased [11C]-deprenyl astrocyte PET binding can be measured 15-20 years before clinical cognitive symp-toms (Rodriguez-Vieitez et al. 2016), thus earlier in AD progression than the increase in plasma GFAP levels (Chiotis et al. 2023). The strong negative association between plasma GFAP and deprenyl PET in autosomal dominant, as well as sporadic AD patients, may most probably be signs for different states or subtypes of as-trogliosis (Chiotis et al. 2023).
As discussed in the paper, the exact underlying mecha-nism by which microglia modulate Aß-induced astrocyte reactivity might still be unknown. Interestingly, astrocytic α7 nicotinic acetylcholine receptors seem to play a cru-cial role, together with soluble Aß, in reactive astro-gliosis as well as in formation of Aß plaques and gluta-mate excitotoxicity (Fontana et al. 2023), which moti-vates further PET studies with new developed PET tracers.
Cliniques Universitaires Saint-Luc and Massachusetts General Hospital
Ferrari-Souza and colleagues offer us an exciting dive into the complex mechanisms linking amyloid and tau pathologies, specifically the role of activated microglia as assessed using both TSPO-PET (C11-PBR28) and sTREM2 CSF concentration. They observed that activated microglia modulated the association between amyloid pathology and tau phosphorylation, tau aggregation, and cognitive impairment. These are important data for secondary prevention trials in A+T- clinically unimpaired individuals, as the risk of (tau/cognitive) progression appears clearly different depending on the state of microglia. They further showed that reactive astrogliosis (GFAP plasma measure) only mediated the association between amyloid and tau when microglia cells were activated. As GFAP associations with tau and cognition depends on microglia state, developing blood-based biomarkers of sTREM2 (or other microglial activation markers) is a critical next step.
BarcelonaBeta Brain Research Center; Hospital del Mar - Barcelona
João Pedro Ferrari-Souza and colleagues present a rigorous and elegant multimodal study addressing a key question: Does microglial reactivity modulate astrocyte reactivity to Aβ in vivo? Using fluid and neuroimaging biomarkers of glial reactivity, the authors show that Aβ relates to astrocytic reactivity (as measured by plasma GFAP) only when microglial reactivity is present (I am sure the authors would agree with me on using this term rather than “microglial activation”). This finding, replicated with both TSPO PET and CSF sTREM2, is further linked to tau phosphorylation/aggregation and cognition, reinforcing the concept of an “amyloid–glia–tau” axis.
This is an excellent study that exemplifies the direction glial biomarker research should take: integrating multimodal biomarkers, validating experimental findings in well-characterized clinical cohorts, and bringing those insights back to the laboratory.
The authors clearly acknowledge the limitations of their work, which are not specific to this study but central to the current discussion in the glial biomarker field (Suárez-Calvet, 2025). Glial and inflammatory responses are dynamic and heterogeneous. Both microglia and astrocytes continuously sense and respond to physiological and pathological changes, with distinct subpopulations exerting protective or detrimental effects depending on context. This inherent complexity limits the interpretation of cross-sectional analyses, though the inclusion of participants across clinical stages in this study helps address part of that challenge. Single readouts such as GFAP, sTREM2, or TSPO capture only specific aspects of this multidimensional process, and understanding what each truly represents remains an important goal. Biomarker–neuropathological correlation studies (Sánchez-Juan et al., 2024) will be key to bridging biological meaning and measurable signals. Future panels of biomarkers and proteomic approaches should aim to resolve multidimensional glial states. Methodological binarization (e.g., “microglia activation positive/negative”) is understandable, but the authors rightly went beyond this approach, performing their analyses not only using binary TSPO or CSF sTREM2 cutoffs but also treating these measures as continuous variables. Distinguishing soluble from aggregated species is also essential. The authors appropriately separate soluble (p-tau217) from aggregated tau; analogous stratification for Aβ (e.g., soluble vs. insoluble) would further strengthen mechanistic interpretation. Notably, in preclinical stages, plasma GFAP appears to mediate the link between CSF Aβ42/40 and Aβ-PET, consistent with an early astrocytic involvement (Pelkmans et al., 2024).
Overall, this carefully executed work positions microglial reactivity as a key modulator of Aβ-related astrocyte responses and downstream tau and cognitive changes, and suggests that early modulation of microglial reactivity may be critical to disease modification. A key question moving forward is how anti-amyloid therapies influence this microglial–astrocytic interplay, and whether their efficacy could be enhanced by concurrent modulation of microglial states.
References:
Pelkmans W, Shekari M, Brugulat-Serrat A, Sánchez-Benavides G, Minguillón C, Fauria K, Molinuevo JL, Grau-Rivera O, González Escalante A, Kollmorgen G, Carboni M, Ashton NJ, Zetterberg H, Blennow K, Suarez-Calvet M, Gispert JD, ALFA study. Astrocyte biomarkers GFAP and YKL-40 mediate early Alzheimer's disease progression. Alzheimers Dement. 2024 Jan;20(1):483-493. Epub 2023 Sep 10 PubMed.
Sánchez-Juan P, Valeriano-Lorenzo E, Ruiz-González A, Pastor AB, Rodrigo Lara H, López-González F, Zea-Sevilla MA, Valentí M, Frades B, Ruiz P, Saiz L, Burgueño-García I, Calero M, Del Ser T, Rábano A. Serum GFAP levels correlate with astrocyte reactivity, post-mortem brain atrophy and neurofibrillary tangles. Brain. 2024 May 3;147(5):1667-1679. PubMed.
Suárez-Calvet M. Fluid and Imaging Markers of Glial Reactivity. Biomarkers of Neurodegenerative Disorders, July 2025 Biomarkers of Neurodegenerative Disorders.
Make a Comment
To make a comment you must login or register.